
Ako uvádza Orel (2021) zmeny vnútri nervovej sústavy (vrátane zmien v mozgu) prebiehajú súbežne so zmenami vonkajšími a sú samozrejme neoddeliteľne spojené so zmenami iných systémov a celého tela. Všetky premeny veľkosti a tvaru embrya a všetkých jeho častí sú podmienené množením, rastom, diferenciáciou buniek, migráciou, i vzájomnou priľnavosťou jednotlivých buniek a bunečných populácií v čase, čo je tiež doprevádzané tiež tvorbou a zmenami priestorového usporiadania buniek a medzibunkového priestoru.
Podľa Orela (2021) tretím týždňom vnútromaternicového života získava zárodok hruškovitý tvar, pretože jeho hlavová časť rastie rýchlejšie a tak je objemnejšia. Vytvorením prechordovej platničky a struny chrbtovej indukujú zhmotnenie ektodermu, ktorý leží nad nimi. Na zadnej dorzálnej strane embrya, tak vzniká neurálna alebo tiež nazývaná ako medulárna platnička, ktorá obsahuje bunky odvodené ektodermu (označujeme ich ako neuroektoderm). Práve neuroektoderm je základovou štruktúrou pre vznik centrálnej nervovej sústavy a mozgu. Neurálna platnička postupne rastie do dĺžky. Na konci tretieho týždňa sa jej vonkajšie okraje vyvyšujú a tvoria neurálne valy, ktoré ohraničujú neurálnu brázdu (ryhu) – pozdĺžne prehĺbenie neurálnej platničky. Neurálne valy sa k sebe postupne približujú až sa nakoniec v strednej časti spoja. Tento proces začína v krčnej chrbtici a postupuje obidvoma smermi (hore a dole). Výsledkom je vznik neurálnej trubice, ktorá sa otvára prostredníctvom predného neuroporu (neuroporus anterior) a dozadu cez zadný neuroporus (neuroporus posterior). Hlavová časť neurálnej trubice, ktorá predstavuje základ mozgu sa rozširuje a záhyby sú na nej dobre viditeľné tri mozgové vačky, rhombencephalon, mesencephalon a prosencephalon. Smerom dole naväzujúca a viditeľne užšia časť neurálnej trubice sa ďalej vyvinie na chrbtovú miechu. Po kompletnom uzavretí oboch neurosporov má centrálny nervový systém podobu uzavretej dutej trubice. Tým je ukončená tvorba neurálnej trubice (neurulácia).
Orel (2021) rovnako tak uvádza, že dutinka vnútri neurálnej trubice sa stane základom pre vznik mozgového komorového systému a centrálneho miechového kanálu, čo budú duté priestory vnútri mozgu a miechy vyplnené mozgomiechovým mokom. Neuroektodermové bunky vonkajšieho okraja neurálnej platničky, ktoré sa k sebe prikladajú a spojujú pri vzniku neurálnej trubice, sa označujú ako neurálna lišta. Neurálna lišta sa postupne začne oddelovať od neurálnej trubice. Jej bunky sa pritom aktívne premiestňujú (migrujú). V oblasti hlavy sa bunky neurálnej lišty oddeľujú z neurálneho valu ešte pred kompletných uzavretím neurálnej trubice. Stanú sa základom pre neuróny a glií v gangliách niektorých hlavových nervov , ale i základom pre vývin melanocytov (buniek produkujúcich tmavohnedý pigment melanín). Kaudálne smerom dole sa vývin a rast gangliovej lišty o niečo oneskoruje a získava segmentové usporiadanie. V oblasti trupu pritom bunky neurálnej lišty využívajú dve možné cesty migrácie:
Ventrálna trasa migrácie smeruje ku zadnej časti smerom dopredu do oblasti brušnej. Neuroektodermové bunky, ktoré využívajú túto cestu sa v nasledujúcom období diferencujú do buniek nervových ganglií miechových spinálnych a vegetatívnych a ďalej do enterického nervového systému, ktoré tvoria dve prepojené pletence autonómnych nervov v stene čriev, popritom dá vznik )pre Schwannove bunky, ktoré vytvárajú obaly neurónov na periférii a časť sa zmení v bunky drene nadobličiek produkujúcich stresové hormóny (epinefrín/adrenalín a norepinefrín/noradrenalín).
Dorzálna trasa migrácie smeruje dozadu do povrchových vrstiev . Neuroektodermové bunky, ktoré tadiaľ putujú, sa následne menia na kožné melanocyty produkujúce hnedý pigment melanín.
Medzi primárne mozgové vačky podľa Orela (2021) patria:
- zadný mozog alebo rhombencephalon obsahuje dve časti: myelencephalon, čo je budúca predĺžená miecha a metencephalon, čo je základ neskoršieho mozočka a Varolova mosta
- stredný mozog alebo mesencephalon
- predný mozog alebo prosencephalon , ktorý sa následne rozdelí na diencephalon, základ medzimozgu, z ktorého vyrastjú očné vačky ako základy sietnice a zrakového nervu a telencephalon , základ mozgových pologúľ
Súčasne sa neurálna trubica ohýba, čím vznikajú dve významné oblasti:
- flexura cervicalis (krčný ohyb) na hranici zadného mozgu a miechy
- flexura cephalica (temenný ohyb) v oblasti stredného mozgu
Zadný mozog a jeho vývin
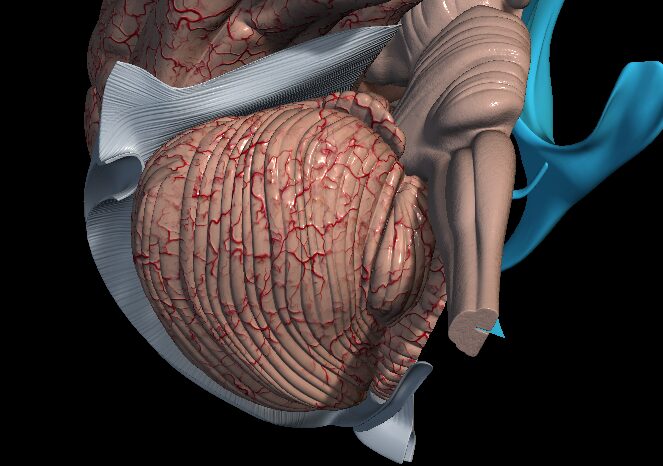
Obrázok č. 1 Rhombencephalon (Zdroj: Anatomy 3D Atlas)
Zadný mozog – rhombencephalon je najnižie uloženým mozgovým vačkom. Členíme ho na myelencephalon a metencephalon. Dáva vznik pre predĺženu miechu, Varlov most a mozoček.
Myelencephalon – bezprostredne naväzuje na časť neurálnej trubice, z ktorej sa vyvinie budúca chrbtová miecha. Tvorí základ pre predĺženú miechu (medula oblongata).
Hlavné časti, ktoré tvoria prvý mozgový vačok, sú dve platničky:
Bazálna platnička – je uložená v strede, nájdeme v nej motorické neuróny ( motoneuróny), ktoré zastupujú oblasť motorickú.
Alárna platnička – je uložená smerom von a obsahuje senzitívne neuróny, reprezentuje teda senzitívnu oblasť.
Nápadným znakom tohto oddielu primitívneho mozgu je otvorenie neurálnej trubice a výrazne oddelenie alárnej platničky žliabkom, ktorý označujeme ako sulcus limitans.
Telá motorických neurónov sú zoskupené do niekoľkých motorických jadier:
Somatomotorické jadrá: sú reprezentované dvomi skupinami, mediálne somatomotorické jadrá, ktoré sú uložené v strede, zatiaľ čo laterálne somatomotorické jadrá ležia smerom von. Obe skupiny jadier budú ovládať priečne pruhované svaly v oblasti hlavy prostredníctvom motorických hlavových nervov (mimické a žuvacie svaly alebo jazyk).
Visceromotorické jadrá: ležia na hranici s alárnou platničkou, teda popri sulcus limitans. Sú určené pre mimovoľné riadenie hladkej svaloviny vnútorných orgánov. Dostanú sa k nim prostredníctvom vegetatívnych nervov. Najmohutnejším vegetatívnym nervov je nervus vagus – blúdivý nerv.
V alárnej platničke nájdeme neuróny senzitívne. I tu sa telá neurónov zoskupujú do jadier, presnejšie senzitívnych jadier:
Senzorické jadrá alárnej platničky bude dostávať informácie o sluchu zo sluchového ústrojenstva a tiež informácie chuťové v chuťových pohárikoch v dutine ústnej.
Somatosenzitívne jadrá sú určené pre spracovanie senzitívnych informácií z oblasti kože a sliznice tváre, dutiny nosnej a ústnej.
Viscerosenzitívne jadrá budú zasobované viscerosenzitívnymi informáciami z vnútorných orgánov (dýchacieho, tráviaceho a kardiovaskulárneho systému).
Na prednej (ventrálnej) strane predĺženej miechy postupne mohutný pár objemných vlákien, ktorém sa bežne označujú ako pyramídy. Obsahujú vlákna kortikospinálne (pyramidové dráhy – tractus corticospinalis), ktoré zostupujú z pyramídových buniek vyvíjajúcej sa motorickej kôry. Táto dráha sa stane základnou cestou riadenia voľnej motoriky, teda vôľou ovládanou hybnosťou. Je treba si uvedomiť, že predĺžená miecha, ktorá vzniká z myelencephala vytvárajú spolu s Varolovým mostom a stredným mozgom, kompaktný útvar, ktorý nazývame truncus cerebri – mozgový kmeň.
Veľmi významná sieť prepojených neurónov, ktorá prechádza celým mozgovým kmeňom sa nazýva retikulárna formácia (formatio reticularis). Tvoria ju vzájomne zložito prepojené neuróny zoskupené do viac ako 50 jadier, ktoré kraniálne (smerom nahor), budú zasahovať až do oblasti medzimozgu. Niektoré jadrá retikulárnej formácie sú špecifické, napríklad kardioexcitačné centrum, kardioinhibičné centrum riadiace srdečnú činnosť , inspiračné (nádychové) a expiračné (výdychové) centrum riadiace dýchanie, centrum reflexu kašľajúceho a kýchajúceho, centrum zvracania, ďalej centrá riadiace reflexy spojené so zrakovým a sluchovým ústrojenstvom. Nie menej významné jadrá mozgového kmeňa sú nešpecifické – plnia prepojovacie a komplexné integračné funkcie. Vo svojej podstate predstavuje retikulárna formácia neobyčajne významnú štruktúru mozgu, ktorá bude hrať kľúčovú rolu v mnohých oblastiach, napríklad v riadení telesných ako je dýchanie, srdečná činnosť, krvný tlak, endokrinné funkcie a pod. a bude významne podielať i na pôvode bdelého stavu. Prispeje tež k riadeniu hybnosti i spracovania rady informácií.
Vzostupný (ascendentný) systém retikulárnej formácie vysiela vlákna do vyšších oblastí mozgu. Vlákna zostupného (descendentného) systému retikulárnej formácie smerujú kaudálne (smerom dole) k neurónom chrbticovej miechy.
Medulla spinalis (chrbticová miecha)
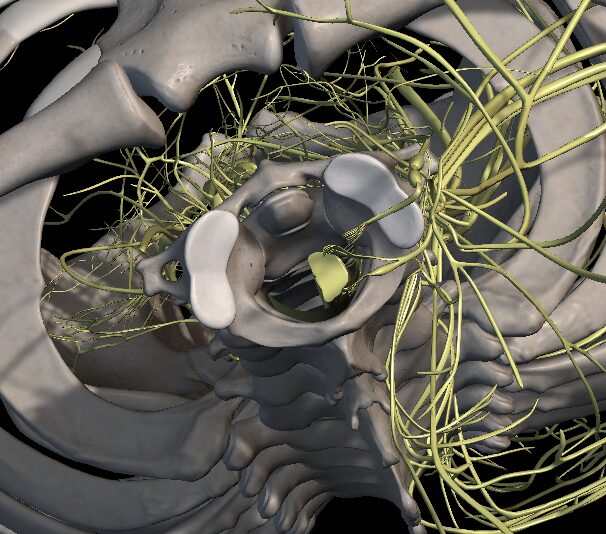
Obrázok č. 2 Medulla spinalis (Zdroj: Anatomy 3D Atlas)
Podľa Kluchovej (2015) je miecha prvá, vývinovo najstaršia časť CNS. Tvorí oblý povrazec nervového tkaniva uložený v chrbticovom kanáli a je obalený blanami. Centrálnu časť miechy predstavuje sivá hmota tvorená telami nervových buniek, okolo ktorej je biela hmota tvorená výbežkami nervových buniek. Do miechy vstupujú senzitívne nervové vlákna a vystupujú z nej motorické vlákna. Senzitívne vlákna vedú podnety všeobecnej citlivosti z periférnych receptorov do miechy. Motorické vlákna prenášajú informácie z miechy na perifériu k výkonnému orgánu. Súčasťou motorických vlákien sú aj autonómne vlákna.
Vonkajší opis
– je uložená v chrbticovom kanáli (canalis vertebralis), tvorí oblý a asi ako prst hrubý povrazec dlhý 42 – 45 cm. Zaberá horné dve dĺžky kanála. Na priečnom reze má oválny tvar a je predozadne spoštená. Miechu obaľujú a chránia tri väzivové blany – povrch miechy kopíruje na cievy bohatá cievnatka, pia mater, ku ktorej zľahka prilieha jemná pavúčnica, arachnoidea. Priestor medzi oboma mäkkými blanami vypĺňa mozgomiechový mok. Tvrdá blana, dura mater, ktorá je lokalizovaná najpovrchnejšie, vytvára okolo miechy a mäkkých obalov vak, saccus durrae matris spinalis, ku ktorému pomerne pevne adheruje pavúčnica. Kraniálne ohraničenie miechy oproti mozgu tvorí veľký záhlavný otvor, foramen magnum, ktorý je vstupom do lebkovej dutiny. Za skutočnú hranicu sa považuje výstup prvého páru krčného nervu (C1), resp. skríženie pyramíd, decussatio pyramidum. Miecha sa kraniálne rozširuje do predĺženej miechy, medulla oblongata, ktorá je tiež súčasťou mozgu. Kaudálne sa zužuje do kužeľovitého útvaru, miechového kužeľa, conus meddularis, ktorý predstavuje ukončenie miechy. Skeletotopicky tak siaha od základnej lebky, basis cranii, až po úroveň druhého a tretieho driekového stavca (L1 – L2). Hrot conus meddularis, siaha u muža do úrovne medzistavcovej platničky medzi L1–L2 a u ženy nižšie, do úrovne tela stavca L2. Úsek chrbticového kanála pod úrovňou stavca L2 už miecha nevypĺňa. Z conus meddularis pokračuje pia mater ako nitkovité vnútorné hraničné vlákno, filum terminale seu piale, ktoré zrastá s periostom druhého krížového stavca (S2). Saccus durae matris fixuje obdobné vonkajšie hraničné vlákno, filum terminale externum seu durale, ktoré dosahuje až ku kostrči. Vnútrom miechy prebieha úzky ústredný kanál, canalis centralis, ktorý sa kraniálne otvára do IV. mozgovej komory a kaudálne končí rozšírením v conus meddularis ako hraničná komôrka, ventriculus terminalis.
Na prednej (ventrálnej) ploche miechy sa v stredovej rovine nachádza hlboká, pozdĺžna predná stredová štrbina, fssura mediana anterior, a po jej stranách, napravo a naľavo, pozdĺžne uložená prodnobočná brázda, sulcus anterolateralis, z ktorej vystupujú koreňové vlákna, fila radicularia, formujúce predné (motorické) miechové korene, radices anteriores.
Na zadnej (dorzálnej) ploche miechy je plytšia, pozdĺžna zadná stredová brázda, sulcus medianus posterior, a po jej stranách zadnobočná brázda, sulcus posterolateralis, je vytvorená zadná prostredná brázda, sulcus intermedius posterior. Predné a zadné miechové korene vznikajú z jemných koreňových vlákien, ktoré sú viditeľné v oboch párových bočných brázdach. Po krátkom priebehu, v tesnej blízkosti medzistavcových otvorov, sa predné a zadné korene spoja a vytvoria miechový nerv, n. spinalis. V priebehu zadného koreňa sa pred miestom spojenia nachádza malá nervová miechová uzlina, ggl. spinale. Úsek miechy, z ktorého vystupuje jeden pár miechových nervov, resp. časť miechy ohraničená predným a zadným miechovým koreňom, sa nazýva miechový segment, segmentum spinale. Miechu tvorí 31 miechových segmentov – 8 krčných (C1 – C8), 12 hrudníkových (Th1 – Th 12), 5 driekových (L1 – L5) a 1 kostrčový segment (Co1). Počnúc segmentom C8, úroveň lokalizácie nižšie uložených segmentov nezodpovedá úrovni príslušných stavcov. Všetky tieto segmenty sú tak lokalizované skeletotopicky o 1 – 3 stavce vyššie, kaudálne siahajú len so úrovne stavca L2, kde sa premieta kostrčový segment (Co1). Predné a zadné korene driekových a krížových miechových nervov, vystupujúce z príslušných segmentov kaudálne od conus medullaris, tak prebiehajú šikmejšie až vertikálne k svojim medzistavcovým otvorom a tvoria zväzok miechových koreňov tvarom pripomínajúcim konský chvost, cauda equina. Miechové nervy L3–Co1 tak nevystupujú z chrbticového kanálu v rovnakej úrovni stavcov ako vznikajú v mieche. Makroskopicky sú na mieche viditeľné dve zhrubnutia vretenovitého tvaru – v krčnej časti miechy – krčné zhrubnutie, intumescentia cervicalis, v rozsahu segmentov C3–Th2 (skeletotopicky C3 – TH1) a v kaudálnej časti miechy driekovokrížové zhrubnutie, intumescentia lumbosacralis, v rozsahu driekových a krížových segmentov L2 – S2 (skeletotopicky Th10 – L1). Zhrubnutia sú tvorené nakopením motorických nervových buniek (motoneurónov) pre svaly hornej a dolnej končatiny, z ktorých vznikajú nervové spleti – plexus brachialis, plexus lumbalis, plexus sacralis.
Vnútorná štruktúra
Miechu tvorí centrálne lokalizovaná sivá hmota, substantia grisea a okolo usporiadaná biela hmota, substantia alba, ktorých štruktúrou je možné skúmať a priečnych rezoch miechou.
Sivá hmota miechy
Na priečnom/tranzverzálnom reze má sivá hmota tvar písmena H a pripomína motýľa. Tvoria ju párové rohy, cornua, ktoré sa do priestoru premietajú ako stĺpce, columnae. Dopredu smeruje predný roh, cornu anterius, do strán, bočný roh, cornu laterate (predĺžená vodorovná čiara v písmene H) dozadu zadný roh, cornu posterius. Rohom zodpovedajú priestorovo usporiadané široké predné stĺpce columne anteriores, malé bočné stĺpce, columnae laterales a štíhle zadné stĺpce, columnae posteriores. Časť sivej hmoty, zona intermedia, spája predný a zadný roh a tvorí tak prostredný stĺpec, columna intermedia, ktorý zahŕňa aj oblasť cornu lateralis. Zona centralis spája pravú a ľavú časť miechového segmentu a tvorí okolo ústredného kanála spredu a zozadu súvislú vrstu sivej hmoty – commisura grisea anterior a commisura grisea posterior.
- Predné stĺpce – columnae anteriores, plnia motorické funkcie (inervujú svaly), obsahujú motorické bunky, ktorých axóny vystupujú predné korene miechy, radices anteriores a vedú vzruchy na perifériu organizmu – k výkonnemu orgánu.
- Zadné stĺpce – columnae posteriores, prijímajú senzitívne informácie, obsahujú senzitívne bunky, na ktorých končí veľká časť axónov pseudounipolárnych buniek miechových ganglií, ktoré tvoria zadné korene miechy, radices posteriores. Cez zadné korene sa prenášajú podnety z periférie organizmu a začínajú tu axóny vzostupných dráh, ktoré pokračujú do vyšsích centier CNS – predĺženej miechy, medzimozgu a koncového mozgu. Zadný roh sa člení na niekoľko častí: základňa/basis, krčok/cervix, hlava/caput a hrot/ cornu posterioris, dosahujúci takmer k povrchu zadnej plochy miechy oproti sulcus posterolateralis, na základe ktorých, sa dá určiť presná lokalizácia senzitívnych neurónov a interneurónov.
- Bočné stĺpce – columnae laterales, plnia viscerálne funkcie, sú vytvorené len v hrudníkovej hornej driekovej a krížovej časti miechy ( v segmentoch Th1 – L2 a S2 – S4). V krčnej časti miechy sa v mieste cornu laterale nachádzajú rôzne roztrúsené jadrá sivej hmoty, ktoré formujú sieťkovitý útvar, formatio reticularis, siahajúce do mozgu.
Biela hmota miechy
Biela hmota obklopuje sivú hmotu. je tvorená myelinizovanými axónmi formujúci systém aferentý (ascendentných), eferentných (descendentných) a asociačných dráh, ktoré prebiehajú homolaterálne, ale aj kontralaterálne cez spojku, commissusa alba. Zväzok axónov tvorí dráhu, tractus, ktorá zabezpečuje spojenie miechy s inou funkčne významnou oblasťou CNS. Biela hmota miechy je rozdelená na tri párové miechové povrazce, funiculi. V povrazcoch prebiehajú aferentné a eferentné nervové dráhy:
Predný povrazec – funiculus anterior: obsahuje aferentné a eferentné dráhy:
- aferentné) predná miechovolôžková dráha, tractus spinothalamicus anterior, je súčasť anterolaterálneho (spinálneho) systému dráh všeobecnej citlivosti, vedie hmatové pocity
- eferentné) predná kôrovomiechová dráha, tractus corticospinalis anterior, je tvorená neskríženými vláknami pyramídovej dráhy (15%), vlákna sa krížia až v úrovni príslušného segmentu v commisura alba a končia na motoneurónoch opačnej strany miechy, vzhľadom k menšiemu počtu vlákien a v porovnaní s mohutnosťou tractus corticospinalis lateralis má táto dráha len veďlajší význam: predsieňovomiechová dráha, tractus vestibulospinalis, sa formuje z predsieňových (vestibulárnych) jadier mozgového kmeňa a prevádza ich vplyv na antigravitačné svalstvo, kontroluje napätie extenzorov končatín: pokrývkovomiechová dráha, tractus tectospinalis,
- je zapojená do hybnosti hlavy a krku v závislosti od prichádzajúcich zrakových vnemov: vsunutá miechová dráha, tractus interstitiospinalis, začína z ncl. interstitialis v strednom mozgu, riadi reflexné pohyby hlavy, krku a hornej časti trupu v závislosti od zrakových a sluchových podnetov.
Zadný povrazec – funiculus posterior, obsahuje hlavnú aferentnú (senzitívnu) dráhu: dráhu jemnej citlivosti, nazývanú aj dráhu zadných povrazcov, tractus spinobulbaris – pre jemnú (epikritickú) rozlišovaciu citlivosť a vibráciu: tvoria ju dva zväzky, štíhly zväzok, fusciculus gracilis – Golli, uložený mediálne a klinovitý zväzok, fasciculus cuneatus – Burdachi, uložený laterálne od štíhleho zväzku.
Bočný povrazec – funiculus lateralis, obsahuje aferentné a eferentné dráhy:
- aferentné)
- bočná miechovolôžková dráha, tractus spinothalamicus lateralis, je súčasť anterloterálneho (spinálneho) systému dráh všeobecnej citlivosti pre bolesť a teplotu, t.j. hrubú (protopatickú) citlivosť, je to dŕaha rýchlej (prenikavej) bolesti,
- miechovosieťová dráha, tractus spinoreticularis, je dráha nepresne lokalizovanej tupej bolesti, dráha pomalej (difúznej) bolesti,
- miechovopokrývková dráha, tractus spinotectalis, je reflexná dráha pre hmatové a polohovocitové informácie v súčasnosti s pohybmi hlavy, krku a očí smerom k hmatovému podnetu,
- miechovoolivová dráha, tractus spinoolivaris, konćí na príslušných jadrách predĺženej miechy a vedie kožné cítenie a propriocepciu zo svalov a šliach oblasti krku,
- predná a zadná miechovomozočková dráha, tractus spinocerebellaris anterior et posterior, sú dráhy proprioceptívnej citlivosti regulujúce svalovú činnosť a napätie a tiež propriorecepciu zo svalov, šliach a kĺbov.
- eferentné)
- bočná kôrovomiechová dráha, tractus corticospinalis lateralis, je hlavná motorická dráha, ktorú tvorí 85% vlákien pyramídovej dráhy krížiacich sa decussatio pyramidum , vlákna končia na motoneurónoch predných rohov miechy,
- červenomiechová dráha, tractus rubrospinalis, začína z ncl. ruber v strednom mozgu, kontroluje napätie flexorov končatín,
- sieťkovomiechová dráha, tractus reticulospinalis, je vývinovo stará motorická dráha, ovplyvňujú autonómne (viscerálne) a motorické funkcie (dýchaciie pohyby, postoj tela a iné)
Funkcie miechy
Miecha zabezpečuje aktívnu reakciu mozgu na zmeny vonkajšieho a vnútorného prostredia organizmu, ktorý na zmeny aktívne reaguje, analyzuje a syntetizuje ich. Spojenie miechy a mozgu je preto nevyhnutné pre koordináciu a integráciu celého tela – koordinačná a integračná funkcia.
Miecha je dôležitým centrom pohybových reflexov – riadi pokojové napätie svalov a jednoduché pohyby. Riadi aj reflexné vyprázdnenie močového mechúra a konečníka – reflexná funkcia. Cez miechu prebiehajú aferentné dráhy, ktoré vedú podnety z receptorov do jednotlivých častí mozgu eferentné dráhy, ktoré vedú informácie z rôznych častí mozgu k výkonným orgánom – prevodová funkcia.
Metencephalon

Obrázok č. 3 Metencephalon (Zdroj: Anatomy 3D Atlas)
– je druhým oddielom zadného mozgu, dáva vznik Varolového mosta, ktorý je súčasťou mozgového kmeňa a rovnako tak aj mozočku.
Mozgový kmeň (truncus encephali) – predĺžená miecha (medulla oblongata), most (pons Varoli) a stredný mozog (mesencephalon).
Ako uvádza Lovásová (2015) mozgový kmeň je časť mozgu uložená v zadnej lebkovej jame. Je pokračovaním miechy a ventrokraniálne prechádza do medzimozgu. Tromi pármi mozočkových stopiek a spája s mozočkom, ktorý je pripojený ku kmeňu zozadu.
Predĺžená miecha (medulla oblongata)
Predĺžená miecha, medulla oblongata seu myelencephalon, je najkaudálnešie uloženou časťou mozgového kmeňa. Je asi 2,5 cm dlhá, 2 cm široká a spája mozog s miechou. Predstavuje rozšírený koniec medulla spinalis a nazýva sa preto aj bulbus medullae spinalis. Kaudálne je ohraničená výstupom prvého krčného miechového nervu, C1 (resp. výstupom koreňových vlákien prvého miechového segmentu), kraniálne siaha po dolný okraj mosta.
Prednou (ventrálnou) plochou je predĺžená miecha uložená na vnútornej spodine lebky, basis cranii interna. Stredom plochy prebieha predná stredová štrbina, fissura mediana anterior, ktorá pokračuje z miechy a kraniálne končí v slepom otvore, foramen caecum. Vpravo a vľavo od štrbiny sa nachádza pozdĺžna vyvvýšenina – pyramída, pyramis. Pyramis laterálne ohraničuje prednobočná brázda, sulcus anterolateralis, z ktorej vystupujú vlákna XII. hlavového nervu, podjazykový nerv, n. hypoglossus. Laterálne od sulcus anterolateralis je uložený asi 1,5 cm dlhý ovoidný útvar oliva, podmienený pod povrchom stočenou platničkou sivej hmoty dolným olivovým jadrom, ncl. olivaris inferior. Kaudálne, na prechode do miechy, sa krížia nervové vlákna kôrovomiechových – pyramídových dráha vytvárajú viditeľné skríženie pyramíd, decussatio pyramidum, ktoré sa tiež považuje za dolnú hranicu medulla oblongata.
Zadnú (dorzálnu) plochu predĺženej miechy prekryvájú hemisféry mozočka. V strede plochy sa nachádza menej výrazná, z miechy pokračujúca, zadná stredová brázda, sulcus medianus posterior, ktorá je kraniálne zakončená malým mostíkom, nazývaným závora, obex. Laterálne od tejto brázdy prebiehajú obojstranne dva hrubé zväzky nervových vlákien: štíhly zväzok, fasciculus gracilis a klinovitý zväzok, fasciculus cuneatus, ktoré sú súčasťou dráh zadných povrazcov, tractus spinobulbaris. Zväzky končia a ďalej sa prepájajú v jadrách – v štíhlom jadre, ncl. gracilis a klinovitom jadre, ncl. cuneatus. Jadrá podmieňujú na zadnej ploche predĺženej miechy pozdĺžne vyklenutia – štíhly hrbček, tuberculum gracile, uložený mediálne a klinovitý hrbček , tuberculum cneatum, uložený laterálne od predchádzajúceho. Laterálne od olivy sa nachádza plytká, zadnobočná brázda, sulcus posterolateralis, z ktorej vystupujú vlákna hlavových nervov IX., X., XI. – jazykovohltanový nerv, n. glossopharyngeus – blúdivý nerv, n. vagus, veďlajší nerv, n. accessorius. Uvedené nervy sú zmiešané, pretože obsahujú senzitívne (somatosenzitívne a viscerosenzitívne), ale aj motorické (somatomotorické a visceromotorické) nervové vlákna. Vystupujú po bokoch predĺženej miechy a nazývajú sa preto bočný zmiešaný systém.
Vnútorná štruktúra – Sivá hmota predĺženej miechy
– je pokračovaním sivej hmoty miechy. Avšak od kompaktnej hmoty v mieche sa líši začným rozčlenením. Nachádzajú sa tu jadrá ncl. gracilis a ncl. cuneatus, v ktorých končia vzostupné senzitívne dráhy zadných povrazcov miechy, tractus spinobulbaris, a začína v nich dráha do lôžka, tractus bulbothalamicus. Preto sa tieto jadrá nazývajú aj jadrá zadných povrazcov. V ich blízkosti sa nachádza aj veďlajšie klinové jadro, ncl. cuneatus accesorius, dolné olivové jadro, ncl. olivaris inferior, pre spojenie s mozočkom a miechou, vedľajšie olivové jadrá a menšie oblúkovité jadrá, ncll. arcuati. Horný úsek zadnej plochy predĺženej miechy tvorí spodinu IV. mozgovej komory. Pod spodinou sa nachádzajú zoskupenia buniek, nazývané jadrá hlavových nervov, ncll. nervorum cranialium, miechové jadro trojklanného nervu, ncl. ambiguus jadro podjazykovéh nervu, ncl. n hypoglossi, zadné jadro blúdivého nervu, ncl. vestibulares, a sluchové (slimákové) jadrá, ncll. cochleares. Stredom sivej hmoty predĺženej miechy prechádza ústredný kanál, ktorý sa rozširuje a ústi do IV. mozgovej komory, ventriculus quartus. Centrálne sa v predĺženej mieche nachádzajú aj jednotlivo roztrúsené nervové bunky, medzi ktorými prebiehajú nervové vlákna a systémy vlákien. Tieto dodávajú na určitých miestach rezu predĺženej mieche sieťový vzhľad a vytvárajú sieťkovitý útvar – retikulárnu formáciu (formatio reticularis).
Biela hmota predĺženej miechy
– obsahuje vzostupné zväzky vlákien – tractus spinothalamicus anterior et lateralis, zoskupené do miechovej stužky, lemniscus spinalis, ku ktorej sa prikladá tractus spitectalis et spinoreticularis. Vnútorné oblúkovité vlákna, fibrae arcuatae internae, ktoré prichádzajú z ncl. gracilis a ncl. cuneatus, sa krížia v útvare nazývanom skríženie stužiek, decussatio lemniscorum a pokračujú kraniálne ako prístredná stužka, lemnicus medialis – zväzok vzostupných senzitívnych dráh, tractus bulbothalamicus. Kranálnou časťou predĺženej miechy prebieha dráha trojklaného nervu, tractus spinalis n. trigemini, ktorá vedie senzitívne podnety do ncl. spinalis n. trigemini. S mozočkom je predĺženej miecha spojená prostredníctvom dolných mozočkových stopiek, pedunculi cerebellares inferiores, ktorými prebieha do mozočka zadná miechovomozočková dráha, tractus spinocerebellaris posterior, dráhy z olivy z ncll. arcuati a z nc. cuneatus accessiorus. Zostupné zväzky vlákien tvorí najmä kôrovomiechová (pyramídová) dráha, tractus corticospinalis, ale aj dráhy z jadier, ale aj dráhy z jadier mosta a stredného mozgu. Vzostupné a zostupné vlákna obsahujú: prístredný pozdĺžny zväzok, fasciculus longuitudinalis medialis a zadný pozdĺžny zväzok, fasciculus longitudinalis posterior.
Varolov most (pons Varoli)
– je mierne rozširujúca časť mozgu, ktorá priamo nadväzuje na predĺženú miechu. Aj Varolov most je rozdelený na bazálnu a alárnu platničku a rovnako tu nájdeme aj motorické a senzitívne neuróny a vieme, že mnohé jadrá predĺženej miechy prechádzajú až sem.
– tvorí nápadný priečny val na prednej ploche mozgového kmeňa. Spája predĺženú miechu a stredný mozog. V strede prednej plochy, ktorá leží na tele záhlavovej kosti a na zadnom okraji tela klinovej kosti, sa nachádza pozdĺžna, spodinová brázda, sulcus basilaris, v ktorej je uložená spodinová tepna, a. basilaris. Brázdu podmieňujú mostom prebiehajúce zostupné zväzky kôrovomiechovej dráhy, tractus corticospinalis, ktoré tvoria miernu klenbu mosta smerom dopredu a majú podobu mohutných valov.
Most tvoria dve vývinovo (ontogeneticky) odlišné časti:
- predná (ventrálna) spodinová časť mosta,pars anterior pontis seu pars basilaris, je vývinovo mladšia, vyvíja sa spolu s rozvojom hemisfér koncového mozgu a obsahuje veľa bielej hmoty, menej je zastúpená sivá hmota
- zadná (dorzálna) pokrývková časť mosta, pars posterior pontis seu pars tegmentalis, je vývinovo staršia, obsahuje bielu hmotu, ale najmä veľké množstvo sivej hmoty
Vnútorná štruktúra mostu – Sivá hmota mostu
V prednej časti mostu, pars anerior pontis, je sivá hmota zoskupená do menších skupín jadier mostu, ncll. pontis, na ktorých končia vlákna prichádzajúce z mozgovej kôry, fibrae corticopontinae.
Zadná časť mostu, pars posterior pontis, nazývaná pokrývka alebo strop mosta, tegmentum pontis, obsahuje jadrá retikulárnej formácie (RF) a jadrá hlavových nervov – jadro odťahujúceho nervu, ncl. n abducentis, jadro tvárového nervu, ncl. n facialis, horné slinné jadro, ncl. salivatorius superior, motorické jadro trojklanného nervu, ncl. motorius n. trigemini a senzitívne jadrá, ncl. principalis (pontinus) et mesencephalicus n. trigemini, predsieňové jadrá, ncll. vestibulares a sluchové jadrá, ncll. cochleares.
Biela hmota mostu
Predná časť mostu obsahuje veľké množstvo bielej hmoty, prevládajú myelinizované, priečne vlákna mostu, fibrae pontis transverae. Sú to axóny buniek ncll. pontis spájajúce most s mozočkom mostovomozočkovou dráhou, tractus pontocerebellaris, ktorá dorzolaterálne podmieňuje mohutné stredné mozočkové stopy, pedunculi cerebellares medii. Pozdĺžne vlákna mosta, fibrae pontis longitudinales, sú súčasťou kôrovomiechovej dráhy, tractus corticospinalis. Tractus corticospinalis spája mozgovú kôru a miechu, vlákna dráhy končia na motoneurónoch predných rohov miechy, fibrae corticospinales, ale aj na motorických dráhach hlavových nervov, fibrae corticonucleares, a v jadrách RF, fibrae corticoreticulares. Na rozhraní prednej a zadnej časti mosta prebieha priečne pásik bielej hmoty, lichobežníkové teleso, corpus trapezoideum. Vlákna telesa sa pridávajú k bočnej stužke, lemnicus lateralis, ktorá je súčasťou sluchovej dráhy. Z medulla oblongata tu pokračujú zväzky nervových vlákien – lemnicus spinalisa lemnicus medialis. Zo senzitívnych jadier, trojklanného nervu vedú dráhy, ku ktorým sa prikladá tractus spinalis n. trigemini a vytvára sa tak trojklanná stužka, lemnicus trigeminalis. V moste prebiehajú aj fasciculus longitudinalis medialis, fasciculus longitudinalis posterior, tractus tectospinalis, rubrospinalis a iné dráhy. Ventrálne, medzi mostom a strednou mozočkovou stopkou, vystupuje z mozgového kmeňa V. hlavový nerv – trojklanný nerv, n. trigeminus. Kaudálne, na rozhraní medulla oblongata a pons, vystupujú v brázde, sulcus bulbopontinus, ďalšie hlavové nervy VI., VII. a VIII. mediálne, bližšie k stredovej rovine – odťahujúci nerv, n. abducens, laterálne od slimákový nerv, n. vestibulocochlearis. N. facialis a n. vestibulocochlearis sú lokalizované v uhle medzi mozočkom, predĺženou miechou a mostom. Uhol sa označuje ako mostovomozočkový uhol, angulus pontocerebellaris.
Kosoštvorcová jama – fossa rhomboidea
IV. mozgová komora je priestor medzi predĺženou miechou, mostom a mozočkom. Zadné plochy medulla oblongata a pons sa podieľajú na tvorbe spodiny IV. mozgovej komory. Spodina má tvar kosoštvorca, od ktorého je odvodený jej názov a nazýva sa kosoštrorcová jama, fossa rhomboidea. Kaudálne sa zužuje do canalis centralis, kraniálne do mozgového mokovodu, aqueductus cerebri. Stredom spodiny prebieha pozdĺžna, stredová brázda, sulcus medianus, ktorú po stranách ohraničuje prístredná vyvýšenina , eminentia medialis. Menej výrazná, pozdĺžne prebiehajúca hraničná brázda, sulcus limitans, sa nachádza laterálne od vyvýšeniny. Fossa rhomboidea sa skladá z troch častí. Dreňové prúžky, striae medullares, prebiehajúce priečne stredom spodiny komory, delia spodinu na: dolnú časť (uloženú kaudálne od prúžkov), prostrednú časť ( jej súčasťou sú uvedené dreňové prúžky ) a hornú časť (uloženú kraniálne od prúžkov). Prúžky súčasne vytvárajú rozhranie medzi oblongata a pons. Sú súčasťou sluchovej dráhy. V prostrednej časti sa priestor IV. mozgovej komory zužuje do bočných zálivov, recessus laterales.
- Dolná časť, pars inferior, je trojuholníkového tvaru a zodpovedá zadnej časti predĺženej miechy. Laterálne ju ohraničuje rozbiehajúce sa pedunculi cerebellares inferiores, ktorými prebiehajú: miechovomozočková dráha, tractus spinocerebellaris posterior, dráhy z olivy – olivomozočková dráha, tractus olivocerebellaris a tiež vonkajšie predné a zadné oblúkovité vlákna, fibrae arcuatae externae anteriores et posteriores, ktoré prebiehajú z oblúkovitých jadier – ncll. arcuati, do mozočka. Z ncl. cuneatus accesorius je do mozočka vedená propriocepcia z horných končatín a hornej časti trupu. Cez dolné mozočkové stopky prebiehajú aj dráhy z ncll.vestibuares do mozočka a z mozočka do jadier v predĺženej mieche. Súčasťou dolnej časti spodiny IV. mozgovej komory sú dve vyklenutia v tvare trojuholníkov – mediálne uložený, svetlejší, kaudálne sa zužujúci trojuholník podjazykového nervu – trigonum n. hypoglossi a laterálne uložený, tmavší trojuholník blúdivého nervu – trigonum n. vagi. Pod trojuholíkmi sú uložené jadrá hlavových nervov XII. a X., ncl. n . hypoglossi et. ncl.posterior n. vagi. Kaudálnu časť trigonum n. vagi ohraničuje malé vaskularizované trojhranné políčko, zadná oblasť, area postrema. Hrot trigonum n. vagi je orientovaný kraniálne do dolnej jamky, fovea inferior, v sulcus limitans.
Medzi dolnými mozočkovými stopkami sa rozprestiera úzka lamela bielej hmoty, dolná dreňová plachta, velum medullare inferius, kaudálne od nej tenká väzivová platnička, cievovkové tkanivo, tela choroidea, s ktorou dolná dreňová plachta zrastá. Tkanivo obsahuje cievovkovú spleť, plexus choroideus. Spleť je dôležitým zdrojom tvorby mozgoviechového moku, ktorý produkuje do IV. mozgovej komory.
- Pod povrchom najširšej, prostrednej časti, pars intermedia, sa nachádzajú rôzne uložené jadrá. Ncl. n. abducenstis spolu s vláknami n. facialis, ktoré jadro kolienkovito obtáčajú, podmieňujú v strede dĺžky eminentia medialis prominujúci útvar, tvárový hrbček, colliculus facialis. Prostredná časť zasahuje do bočných zálivov, v spodine ktorých ležia skupiny predsieňových a sluchových jadier, ncll. vestibulares et cochleares. Predsieňové jadrá podmieňujú zaoblenú vyvýšeninu na začiatku bočných zálivov, predsieňovú oblasť, area vestibulares. Sluchové jadrá ležia v zálivoch dorzolaterálne a podmieňujú vyvýšeninu, sluchový hrbček, tuberculum acusticum.
- Horná časť spodiny IV. mozgovej komory, pars superior, tiež v tvare trojuholníka, zodpovedá zadnej časti mosta. Laterálne ju ohraničujú horné mozočkové stopy, pedunculi cerebellares superiores. Stopky spájajú mozoček so stredným mozgom. Cez stopky prebieha do kôry mozočka predná miechovomozočková dráha, tractus spinocerebellaris anterior a z mozočka dráhy ku štruktúram mozgového kmeňa a lôžka. Medzi stopkami sa rozprestiera horná dreňová plachta, velum medullare superius. Na spodine komory, v sulcus limitans, je lokalizovaná horná jamka, fovea superior, a v jej blízkosti sa laterálne nachádza zoskupenie pigmentových buniek, belasé miesto, locus cauruleus, ktoré tvorí noradrenalín.
Mozoček (Cerebellum)
Mozoček je druhou štruktúrou, ktorá sa vedľa Varolovho mosta vyvíja z mezencephala. Časti alárnych platničiek uložených v zadnej a vonkajej časti metencephala sú silné a časom ďalej výrazne mohutné. Vytvárajú valy, ktoré rastú a tvarujú mozočkovú platničku. Už v 12. týždni vnútromaternicového života na nej môžeme rozlíšiť nepárovú strednú časť (budúci mozočkový červ – vermis cerebelli) a smerom do von uložené základy dvoch mozočkových pologúľ (mozočkových hemisfér). V ďalšom vývine hmota mozočku ( najmä mozočkových hemisfér) mohutnie, až celkom prekryje oblasť predĺženej miechy a Varolovho mosta. Na povrchu mozočka sa postupne objavujú ryhy, ktoré mu dávajú charakteristický a známy vzhľad. Mozočkovú platničku tvoria tri bunečné vrstvy, diferencuje sa vnútorná a vonkajšia germinálna (zarodočná) vrstva. Množením a ďalším vývinom neuroblastov v tejto oblasti vznikajú jednotlivé bunky mozočka. Podstatnou súčasťou vývinu neurónov mozočku je ich migrácia. Napríklad Purkyňove bunky mozočkovej kôry majú pôvod vo vnútornej germinálnej vrstve, pričom sa aktívne premiestňujú smerom k povrchu. Neuroblasty vonkajšej germinálnej vrstvy sa diferencujú na bunky hviezdicové a košíčkové. Neuroblasty vnútornej germinálnej vrstvy, ktoré neputujú, davajú vznik pre mozočkové jadrá uložené v bielej hmote v hĺbke mozočku. Veľmi dôležitou súčasťou mozočka sú aj podporné bunky, ktoré pochádzajú ako z vonkajšej, tak z vnútornej germinálnej vrstvy.
Štruktúra mozočka odráža jeho vývin, v priebehu ktorého postupne vznikajú a rozvíjajú sa tri mozočkové oblasti – archicerebellum, palleocerebellum a konečne neocerebellum:
Archicerebellum (vestibulárny mozoček) – predstavuje bazálnu oblasť mozočka, ktorá vzniká ako prvá. Jej funkcia je spojená s rovnovážnym ústrojenstvom. Podieľa sa na udržiavaní vzpriameného postoja a rovnováhy.
Paleocerebellum (spinálny mozoček) – zahrňuje mozočkového červa a prilahlé (paravermálne) oblasti mozočkových pologúľ. Zpracuváva najmä dáta z končatín a významne sa podieľa ma regulácií svalového napätia.
Neocerebellum ( kôrový mozoček) – je vývinovo najmladšia časť mozočku, reprezentujú ho vonkajšie časti mozočkových pologúľ. Spolupodieľa na riadení precíznosti, jemnosti a koordinácie pohybu. Vstupuje tiež do prežívania, správania a procesu učenia.
Zrelá mozočková kôra je členená do troch vrtiev, ktoré sa postupne vyvíjajú. Obsahujú rôzne typy neurónov, ktoré sú vzájomne prepojené, a ktoré vysieľajú a prijímajú vlákna do/z iných oblastí mozgu a miechy:
Stratum moleculare – je vrstvou uloženou najvrchnejšie, obsahuje hviezdicové a košičkové bunky, ktoré sú v intenzívnom kontakte s Purkyňovými bunkami.
Stratum gangliosum – (stredná vrstva mozočkovej kôry) je tvorená vrstvou veľkých buniek Purkyňových.
Stratum granulosum – je najhlbšou vrstvou mozočkovej kôry, obsahuje bunky zrnité a Golgiho bunky.
Mozoček je bohato prepojený s inými časťami mozgu. Deje sa tak prostredníctvom (aferentných) a odstredivých (eferentných) nervových vlákien, ktorá prebieha tromi mozočkovými stonkami (hornými, strednými a dolnými):
Aferentné vlákna smerujú do mozočkové kôry a privádzajú do nej informácie z chrbticovej miechy, mozgového kmeňa, mozgovej kôry a podkôrových oblastí. Označujú sa ako miechové a šplhavé vlákna.
Eferentné vlákna vystupujú z mozočkových jadier (zhlukov mozočkových neurónov) uložených v hĺbke mozočku a tiež z mozočkovej kôry. Ich výstupy smerujú najmä do mozgového kmeňa a thalamu ( a ich prostredníctvom do ďalších oblastí mozgu chrbticovej miechy).
Ako uvádza Boleková (2015) mozoček vytvára spolu s mostom zadný mozog. Váži priemerne 130g. Je uložený nad mozgovým kmeňom.
Vonkajší opis
Mozoček sa skladá z dvoch hemisfér, hemispheriae cerebelli, ktoré sú v stredovej rovine spojené mozočkovým červom,, vermis cerebelli. Mozoček je uložený v zadnej lebečnej jame a od záhlavového laloka koncového mozgu ho oddeľuje tvrá blana, mozočkový šiator, tentorium cerebelli. Dolná plocha mozočka tvorí strop IV. mozgovej komory. Povrch mozočka je rozčlenený na množstvo paralelne prebiehajúcich mozočkových listov, folia cerebelli, ktoré sú navzájom oddelené mozočkovými štrbinami, fissurae cerebelli. Mozoček je spojený s mozgovým kmeňom prostredníctvom troch párov mozočkových stopiek, pedunculi cerebellares:
- horné mozočkové stopy, pedunculli cerebellares superiores, spájajú mozoček so stredným mozgom
- stredné mozočkové stopy, pedunculi cerebellares medii, spájajú mozoček s mostom
- dolné mozočkove stopy, pedunculi cerebellares inferiores, spájajú mozoček s predĺženou miechou
Z morfologického hľadiska sa mozoček delí na tri laloky:
- predný lalok, lobus cerebelli anterior
- zadný lalok, lobus cerebelli posterior
- chumáčikovouzlovitý lalok, lobus flocculonodularis
Jednotlivým častiam sa vermis zodpovedajú presné časti na hemisférach. Lobus cerebelli anterior et posterior navzájom oddeľuje prvá štrbina, fissura prima, medzi lobus cerebelli posterior et fluccolonodularis sa nachádza zadnobočná štrbina, fissura psterolateralis. Vodorovná štrbina, fissura horizontalis, oddeľuje hornú plochu mozočka od dolnej.
Vnútorná štruktúra mozočka
- mozočková kôra, cortex cerebelli, pokrýva povrch mozočka a nemá schopnosť uchovávať pamäťové informácie
- dreňové teleso mozočka, corpus medullare cerebelli, je vnútri mozočka a rozvetvuje sa smerom k povrchu do sivej hmoty tak, že na mediánnom reze mozočkových červom vytvára útvar v tvare rozkonáreného stromu s názvom strom života, arbor vitae, bielu hmotu mozočka tvoria aferentné a eferentné mozočkové dráhy
- vnútri bielej hmoty mozočka sa nachádzajú zhluky sivej hmoty, ktoré tvoria párové mozočkové jadrá, ncll. cerebelli
Sivá hmota mozočka
Cortex cerebelli sa skladá z troch vrstiev:
- vonkajšia vrstva, stratum moleculare – obsahuje hviezdicovité a košíčkovité bunky, ktoré sú roztrúsené medzi bohato rozvetvenými dendritmi Purkyňových buniek,
- stredná vrstva, stratum purkinjense – obsahuje veľké Purkyňove bunky, ktorých dendrity siahajú do stratum moleculare, axóny Purkyňových buniek končia v jadrách mozočka a vo vestibulárnych jadrách,
- vnútorná vrstva, stratum granulosum – obsahuje granulárne bunky, ktorých axóny končia v stratum moleculare a Golgiho bunky, ktorých dendrity vstupujú do všetkých vrstiev kôry
Do mozočka prichádzajú dva druhy aferentných nervových vlákien.Šplhavé vlákna prechádzajú cez všetky vrstvy do stratum moleculare a končia na dendritoch Purkyňových buniek. Machové vlákna sa končia na granulárnych bunkách stratum granulosum.
Mozočkové jadrá
V bielej hmote mozočka sa nachádzajú 4 typy párových mozočkových jadier, ncll. cerebelli:
- ozubené jadro, ncl. dentatus
- zátkovité jadro, ncl. emboliformis
- guľovité jadrá, ncll. globosi
- jadrá povaly, ncll. fastigii
Biela hmota mozočka
Aferentné dráhy mozočka obchádzajú mozočkové jadrá, idú priamo do kôry mozočka a privádzajú informácie z miechy a častí mozgu o polohe a pohybe tela o svalovom napätí:
- predná miechovomozočková dráha, tractus spinocerebellaris anterior – začína v zadnom rohu miechy, v ncl. thoracicus posterior a prebieha cez pedunculus cerebellaris superior do kôry mozočka
- zadná miechovomozočková dráha, tractus spinocerebellaris posterior – z ncl. thoracicus posterior miechy vedie cez pedunculus cerebellaris inferior do kôry mozočka
- klinovomozočková dráha, tractus cuneocerebellaris – z ncl. cuneatus accessorius predĺženej miechy cez peduncules cerebellaris inferior do kôry mozočka
- olivomozočková dráha, tractus olivocerebellaris – z ncll. olivares predĺženej miechy cez pedunculus cerebellaris inferior do kôry mozočka
- predsieňovomozočková dráha, tractus vestibulocerebellaris – z ncll. vestibulares mozgového kmeňa cez peduncules cerebellaris inferior do nosulus a flocculus
- mostovomozočková dráha, tractus pontocerebellaris – z ncll. pontis cez pedunculus cerebellaris medius do kôry mozočka
Eferentné dráhy začínajú v mozočkovej kôre na axónoch Purkyňových buniek, idú do mozočkových jadier – k motorickým centrám mozgového kmeňa, vestibulárnym jadrám, k jadrám RF a lôžka, podieľajú sa na koordinácií pohybov a regulácií svalového napätia:
- mozočkovopredsieňová dráha, tractus cerebellovestibularis – z ncll. fastigii cez pedunculus cerebellaris inferior do ncll. vestibulares
- mozočkovosieťkovitá dráha – tractus cerebelloreticularis – z ncl. dentatus, ncl. emboliformis a z ncll. globosi cez pedunculus cerebellaris superior k RF
- mozočkočervená dráha, tractus cerebellorubralis – z ncl. dentatus, ncl. emboliformis a ncll. globosi cez pedunculus cerebellaris superior do ncl. ruber stredného mosta
- mozočkovolôžková dráha, tractus cerebellothalamicus – z ncl. dentatus, ncl. emboliformis a z ncll. globosi cez pedunculus cerebellaris superior do ventrálnych jadier lôžka, ncll. ventrales thalami
Funkcia mozočka
Mozoček reguluje svalové napätie a zabezpečuje správnu koordináciu pohybov. Má spojenia s mozgovou kôrou a bazálnymi gangliami, s mozgovým kmeňom aj s miechou. Hlavný zdroj informácií o polohe a pohybe hlavy a končatín prichádza do mozočka z vestibulárnych a miechových dráh.
Mozoček sa zúčastňuje na riadení nielen úmyselných, chcených pohybov, ale aj na riadení mimovôľových, reflexných pohybov. Mozočková kôra reguluje svalové napätie a zabezpečuje koordináciu svalových pohybov, riadi udržiavanie rovnováhy tela pri vzpriamenej polohe. Mozoček koordinuje aj úmyselné a rýchle pohyby. Rýchly sled impulzov z mozočka ku svalom tvorí základ regulácie rýchlo sa striedajúcich opakovaných pohybov. Mozočkové dráhy myelinizujú okolo 5.–6. mesiaca života, kedy u dieťaťa nastupuje koordinovaná pohybová aktivita.
Stredný mozog a jeho vývin
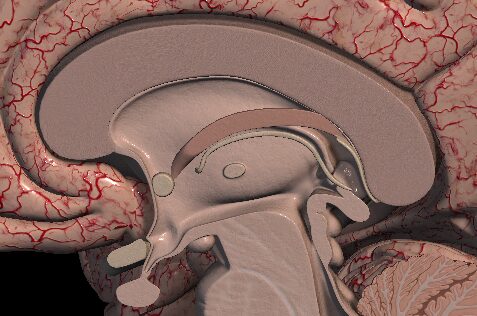
Obrázok č. 4 Mesencephalon (Zdroj: Anatomy 3D Atlas)
Podľa Orela (2021) v bazálnej platničke na spodnej strane stredného mozgu (mesencephala) sa postupne diferencujú motorické neuróny, ktorých zoskupením vzniknú jadrá niektorých hlavových nervov. Pod bazálnou platničkou vznikajú povrazcovité útvary (crura cerebri), ktorými prechádzajú axóny z vyšších oblastí mozgu do predĺženej a chrbticovej miechy. Alárne platničky na hornej časti )stredného mozgu majú spočiatku podobu dvoch hrboľkov (corpora bigemina) oddelené žliabkom uloženým v strede. Následne sa oba hrboly rozdelia priamym žliabkom, čím vznikne charakteristické štvorhrbolie skladajúce sa z párových colliculi superiores, uložených hore, a párových colliculi inferiores nachádzajúcich sa dole. Rovnako vo vývine stredného mozgu hrá nezastúpiteľnú úlohu migrácia neuroblastov, ktorá smeruje z hĺbky k povrchu a prebieha v niekoľkých vlnách. Vďaka tomu vzniká vrstevnaté usporiadanie, zrejme najmä v hornej časti štvorhrbolia.
Lovásová (2015) spomína, že mesencephalon tvorí krátku časť mozgového kmeňa (asi 1,5 – 2 cm). Kaudálne je spojený s mostom, kraniálne s medzimozgom. Zadná (dorzálna) plocha a bočné plochy stredného mozgu sú prekryté hemisférami koncového mozgu. Len predná (ventrálna) plocha je viditeľná na spodine mozgu, basis cerebri, ako dva mohutné rozostupujúce sa valy – mozgové ramená, crura cerebri. Na priereze stredným mozgom vidieť úzky kanál, mozgový mokovod, aqueductus cerebri (mesencephali), ktorý spája III. a IV. mozgovú komoru. Rovina preložená týmto kanálom delí stredný mozog na prednú a zadnú časť.
- Predná (ventrálna) časť – mozgová stopka, pedunculus cerebri
- Zadná (dorzálna) časť – pokrývka stredného mozgu, strecha, tectum mesencephali
Vrstva čiernosivej hmoty, čierna hmota, substantia nigra, ktorá obsahuje veľa pigmentových buniek, delí pedunculus cerebri –
- prednú (ventrálnu) časť – mozgové ramená, crura cerebri
- zadnú (dorzálnu) časť – pokrývku, strop, tegmentum mesencephali
Tectum mesencephali je tvorené platničkou pokrývky, lamina tecti, na ktorej sa nachádzajú dva páry hrbčekov – vyššie a užšie horné hrbčeky, colliculi superiores a nižšie a širšie dolné hrbčeky, colliculi inferiores.
Jadrá horných hrbčekov, ncll. colliculi superiores, sú podkôrovými zrakovými centrami riadiacimi koordinované pohyby očnej gule, hlavy a následne celého tela – sú centrom opticko-motorických reflexov. Zodpovedajú za reflexný pohyb očí a hlavy za zdrojom svetla a za obranný reflex – rýchle otočenie hlavy vyvolané svetelným podráždením sietnice. Vpredu, pred colliculi superiores, sa nachádza predpokrývková oblasť, area pretectalis, ktorej jadrá, ncll. pretectales, sú reflexnými centrami pupilárnej kontrakcie – zúženia zremice, miosis a rozšírenie zrenice, mydriasis.
Jadrá dolných hrbčekov, ncll. colliculi inferiores, sú prepojovacou stanicou sluchovej dráhy – sú centrom akusticko-motorických reflexov. Pod colliculi inferiores, v stredovej rovine, je uzdička hornej dreňovej plachty, frenulum veli medullaris superioris, pre úpon stroku IV. mozgovej komory. Po stranách uzdičky vystupuje IV. hlavový nerv – kladkový nerv, n. trochlearis, ktorý ako jediný a najtenší z hlavových nervov vystupuje z dorzálnej plochy mozgového kmeňa. Prostredníctvom oblých ventrolaterálne prebiehajúcich valov, ramien dolného a horného hrbčeka, brachium colliculi superioris a brachium colliculi inferioris, sa spájajú horné a dolné hrbčeky s časťou medzimozgu – so zálôžkom metathalamus.
Vnútorná štruktúra – Sivá hmota stredného mozgu
Tectum mesencephali je tvorené vrstvičkami sivej hmoty v culliculi superiores, v ktorých sú obsiahnuté jadrá horných hrbčekov, ncll. colliculi superiores a väčším jadrom dolného hrbčeka, ncl. colliculi inferioris, ktoré je lokalizované v colliculus inferior. Tegmentum mesencephali obsahuje väčšie množstvo sivej hmoty – sivú hmotu okolo aqueductus cerebri, substantia grisea centralis, červené jadro, ncl. ruber, substantia nigra, jadrá formatio reticularis, jadrá hlavových nervov III. a IV. – jadro okohybeho nervu, ncl. n. oculomotorii, vedľajšie (autonómne) jadro okohybného nervu, ncl. accessorius (autonomicus) n. oculomotorii, nazývame tiež Edinger–Westphalovo jadro a jadro kladkového nervu, ncl. n. trochlearis. Nucleus ruber je veľký podlhovastý až guľovitý útvar sivej hmoty uložený v centrálnej časti tegmentum mesencephali. Na reze je viditeľný ako naružovelé oválne jadro. Je miestom prepojenia dráh zúčastňujúcich sa na kontrole motoriky. Substantia nigra je platnička čiernosivej hmoty, zloženej kompaktnej tmavšej ústrednej časti, pars compacta a pigmentovej svetlejšej sieťkovotej časti, pars reticularis. Pars reticularis obsahuje prevažne žltohnedý pigment lipofuscín. Pars compacta patrí k monoaminergickému systému mozgového kmeňa, tvoria ju husto nakopené nervové bunky obsahujúce čierny pigment neuromelanín, vedľajší produkt tvorby neuromediátora – dopamínu. Funkčne je substantia nigra zapojená do kontroly systémov motoriky a má prepojenie s bazálnymi gangliami. Ostatné jadrá lokalizované v tegmentum mesencephali: stredové jadro, ncl. medianus (Perliovo jadro), je menšie jadro zabezpečujúce konvergentné pohyby očí, vsunuté jadro, ncl. interstitialis (Cajalovo jadro) a ncl. Darkschewitschi (Darkševičovo jadro), všetky dostávajú prívody z colliculi superiores, ncll. vestibulares , z globus pallidus a z kôry, vysielajú vlákna do fasciculus longitudinalis medialis, čím je zabezpečená koordinácia pohybov hlavy a krku s pohybmi očí v závislosti na orgáne rovnováhy.
Biela hmota stredného mozgu
Tectum mesencephali tvoria: vrstvičky bielej hmoty v colliculi superiores (časť zrakových vlákien), koncový úsek lemnicus lateralis, začínajú tu dráhy smerujúce do kaudálnejších častí. Biela hmota v tegmentum mesencephali je tvorená zväzkami senzitívnych dráh – lemniscus lateralis, motorickými dráhami z kôry, z jadier RF, z ncl. ruber, prebiehatu fasciculus longitudinalis posterior. V tegmentum mesencephali začína zväzok vlákien fasciculus longitudinalis medialis, ktorý prebieha celou dĺžkou mozgového kmeňa a spája Cajalovo, Darkševičovo jadro, okohybné jadrá, predsieňové jadrá a motoneuróny miechy pre svaly krku. Crura cerebri obsahujú niekoľko systémov vlákien – zväzky zostupných motorických dráh, tractus corticospinalis. Vlákna prichádzajú z mozgovej kôry cestou vnútorného púzdra, capsula interna a prechádzajú cez crura cerebri ako fibrae corticopontinae, corticonucleares et capsula interna a prechádzajú cez crura cerebri ako fibrae corticopontinae, corticonucleares et corticospinales. Časť vlákien sa prepája v jadrách mozgového kmeňa, časť pokračuje kaudálne mozgovým kmeňom do miechy. Na spodine mozgu sa medzi rozbiehajúcimi sa crura cerebri nachádza medzistopková jama,fossa interpeduncularis. Spodinu jamy tvorí nervové tkanivo s ostrovčekmi pre cievy – medzistopková (zadná) dierkovaná hmota, substancia perforata interpeduncularis seu posterior. V jej bočnom okraji vystupuje III. hlavový nerv – okohybný nerv, n. oculomotorius. Pod fossa interpeduncularis sa nachádza malá skupina nervových buniek, medzistopkové jadro, ncl. interpeduncularis. Jadro je miestom prepojenia čuchových reflexných dráh a je zapojené aj v okruhoch limbického systému.
Jadrá hlavových nervov (nuclei nervorum cranialium)
– sú lokalizované v mozgovom kmeni, prevažne pod fossa rhomboidea. Len jadrá III. a IV. hlavového nervu sú uložené v strednom mozgu a jadro XI. hlavového nervu zasahuje kaudálne do krčnej časti miechy. Podľa toho, či z jadier vystupujú motorické nervové vlákna, alebo v nich končia senzitívne nervové vlákna, sa označujú ako nuclei origins alebo nuclei terminationis.
Ncll. originis funkčne zodpovedajú skupinám motoneurónov v predných a bočných stĺpcoch miechy. Ncll. terminationis sú lokalizované laterálne od predchádzajúcich a zodpovedajú senzitívnym neurónom – jadrám zadných stĺpcov miechy a jadrám zadných povrazcov.
Jadrá tvorí 8 funkčných komponentov: motorické a senzitívne, somatomotorické a somatosenzitívne, visceromotorické a viscerosenzitívne, všeobecné a špeciálne. Jadrá sú usporiadané do pozdĺžnych pruhov tak, že somatomotorické a visceromotorické jadrá ležia mediálne od sulcus limitans, viscerosenzitívne a somatosenzitívne jadrá sú uložené laterálne od tej brázdy azaberajú aj oblasť spodiny v recessus ôateralis (area vestibularis) ventriculi quarti. Autonómne jadrá hlavových nervov sú uložené uprostred medzi motorickými ncll. originis a senzitívnymi ncll. terminationis – visceromotorické jadrá sú uložené v blízkosti ncll. originis viscerosenzitívne v blízkosti ncll. terminationis.
Somatomotorické jadrá
- Všeobecné somatomotorické jadrá – ncl. hypoglosi
- Špeciálne somatomotorické jadrá – ncl. n. abducentis, ncl. n. trochlearis, ncl. n. oculomotorii
Uvedené jadrá tvoria dorzomediálny rad motorických jadier, sú uložené pod povrchom spodiny IV. mozgovej komory a v strednom mozgu a sú usporiadané v mozgovom kmeni vzostupne. Neuróny týchto jadier predstavujú motorické neuróny, ktoré inervujú priečne pruhované svaly jazyka a okohybné svaly. Somatomotorické jadrá sú zapojené do systémov motoriky a reflexných oblúkov.
Visceromotorické jadrá
Všeobecné visceromotorické jadrá sú tvorené pregangliovými neurónmi parasympatika. K výkonnému orgánu sa vždy prepájajú v parasympatikovom gangliu na postgangliový neurón. Sú to autonómne parasympatikové jadrá zapojené do viscerálnych miechových reflexov. Zabezpečujú inerváciu hladkej svaloviny očnej gule, dýchacieho, tráviaceho systému, priečne pruhovanej svaloviny srdca, veľkých slinných žliaz, malých slinných žliaz a slznej žľazy. Visceromotorické jadrá tvoria dorzolaterálny rad motorických jadier. Patria sem: ncl. accessorius (autonomicus) n . oculomotorii – pre inerváciu zvierača zrenice a vráskovcového svalu (zúženie zrenice a akomodácia oka), ncl. posterior n. vagi – pre inerváciu dýchacieho, tráviaceho systému a srdca, ncl. salivatorius inferior, nazývané aj ako zadné jadro jazykovohlatanového nervu, ncl. posterior n. glossopharyngei – pre inerváciu príušnej žľazy, drobných žliaz sliznice líc, nosohltana, sluchovej trunice a sliznice stredoušnej dutiny, ncl. salivatorius superior, nazývané aj zadné jadro tvárového nervu, ncl. posterior n . facialis – pre slznú žľazu, podjazykovú žľazu, podsánkovú žľazu a drobné žliazky v sliznice ústnej a nosovej dutiny.
Špeciálne visceromotorické (branchiomotorické) jadrá tvoria skupinu pôvodne všeobecných visceromotorických jadier, ktoré však v definitívnom usporiadaní, vo vzťahu ku svalom ovládaných vôľou, fungujú ako somatomotorické a nazývajú sa preto špeciálne jadrá. Tieto sú uložené hlbšie pod spodinou fossa rhomboidea a sú z nich inervované priečne pruhované svaly pochádzajúce z materiálu žiabrových oblúkov. Patria sem ncl. ambiguus (nazývaný tiež ako ncl. originis n. glossopharyngei, n. vagi et n. accessori), ncl. n. facialis a nlc. motorius n. trigemini). Prostredíctvom IX. a X. hlavového nervu sú z príslušného jadra inervované svaly podnebia, hltana, hrtana, pažeráka. Z motorického jadra je zabezpečená inervácia žuvacích svalov a niektorých svalov krku.
Viscerosenzitívne jadrá
Skupinu viscerosenzitívnych jadier tvorí len jediné jadro, ktoré prijíma všetky viscerálne podnety. Aferentné vlákna z ganglií IX. a X. resp. VII. hlavového nervu vedú viscerálnu senzitivitu z dýchacieho a tráviaceho systému, zo srdca, z baroreceptorov a chemoreceptorov cievneho systému a tvoria v mozgovom kmeni zväzok descendentných kolaterál, osamotenú dráhu, tractus solitarius, okolo ktorej sú zoskupené viaceré menšie jadrá, tvoriace jednotné jadro osamotenej dráhy, ncl. tractus solitarii – ncl. splitarius. V tomto jadre končia všeobecné viscerosenzitívne vlákna IX. a X. hlavového nervu. Jadrá v strednej časti osamoteného jadra, ncl. solitarius, sa podieľajú na formovaní kariovaskulárneho centra, jadrá v kaudálnej časti formujú dýchacie centrum a riadia reflexy spojené s dýchaním.
Všeobecné viscerosenzitívne jadro predstavuje stredná a kauzálna časť jadra a nazýva sa spojkové jadro, ncl. commissuralis.
Skupinu špeciálnych viscerosenzitívnych jadier tvorí kraniálna časť, ncl. solitarius prostredníctvom aferentných vlákien z ganglií VII. hlavového nervu a vyššie uvedených hlavových nervov IX. a X. dostala chuťové podnety z chuťových receptorov jazyka, podnebia a hrtanovej príchlopky, nazýva sa chuťové jadro, ncl. gustatorius.
Somatosenzitívne jadrá
Skupina somatosenzitívnych jadier prijíma informácie z receptorov všeobecnej citlivosti a zo zmyslových orgánov. Z receptorov všeobecnej citlivosti prijímajú senzitívne podnety jadrá n, trigeminus, ktoré sú lokalizované laterálne od sulcus limitans a ncl. solitarius. Somatosenzitívne vlákna, ktoré prebiehajú v iných hlavových nervoch (v VII., IX., a X) sa po vstupe do mozgového kmeňa od týchto nervov oddelia a vstupujú do spoločného zväzku s vláknami V – tractus spinalis nervi trigemini.
Všeobecné somatosenzitívne jadrá – ncl. spinalis n. trigemini, ncl. principalis (pontinus) n.trigemini a ncl. mesencephalicus n. trigemini. Stĺpce týchto jadier sa rozprestiera v celej dĺžke mozgového kmeňa. Jadrá prijímajú cestou aferentných vlákien ganglií hlavových nervov V.,VII., IX. a X. impulzy z receptorov všeobecnej citlivosti: podnety bolesti, teploty, dotyku, tlaku, vibrácie a propriorecepcie z oblasti hlavy. Jadrá zodpovedajú funkčne senzitívnym jadrám zadných rohov miechy – ncl. spinalis n. trigemini et ncl. mesencephalicus n. trigemini a jadrám zadných povrazcov – ncl. principalis n. trigemini. Prijímajú však senzitivitu z oblasti hlavy.
Špeciálne somatosenzitívne (senzorické) jadrá zastupujú dve skupiny jadier, ktoré sú uložené najlaterálnejšie, pod spodinou recessus lateralis. Patria sem: ncll. cochleares, ktoré tvoria dve jadrá a ncll. vestibulares, ktoré sú rozdelené do štyroch samostatných jadier. Uvedené senzorické jadrá prijímajú podnety zo zmyslových orgánov – zo sluchového a predsieňového orgánu (systému) vnútorného ucha. Jadrá sú zapojené do systémov senzorických dráh – sluchovej a predieňovej (vestibulárnej).
Retikulárna formácia – formatio reticularis
– spojovacie a koordinačné ústredie, ktoré zaisťuje mnohé prepojenia vnútri CNS. Nachádzajú sa v ňom dôležité centrá pre riadenie vitálnych funkcií, je centrom dôležitých reflexov – obživných a obranných. V RF sa nachádzajú koordinačné centrá pre reguláciu dýchania, vazomotoriku, činnosť srdca a pneumotaktické centrum. RF plní mnohé aktivačné a inhibičné funkcie. Pôsobí budivo na mozgovú kôru a podieľa sa tak na kontrole cyklu bdenie – spánok a vytváraní stavu bdenia. RF je zapojená do procesu pomalej nepresne lokalizovanej difúznej bolesti.
Sieťkovitý útvar, nazývaný aj retikulárna formácia, formatio reticularis, je vývinovo stará, ale z hľadiska funkcie veľmi dôležitá časť CNS. Rozprestiera sa od hornej, krčnej časti miechy až po stredný mozog. V rozsahu predĺženej miechy dosahuje RF takmer k spodine IV. mozgovej komory, v moste je lokalizovaná centrálne – v tegmentum pontis a v strednom mozgu tiež centrálne – v tegmentum mesencephali. RF tvorí súbor neurónov medzi motorickými a snzitívnymi jadrami hlavových nervov. Tento útvar v mozgovom kmeni sa člení do väčších skupín retikulárnych jadier, ncll. reticulares, usporiadaných do troch pozdĺžnych pruhov, nepárneho stredového rafeálneho systému a dvoch párových systémov, mediálneho a laterálneho. RF získava senzitívne podnety z ďalších častí mozgu a miechy a prepája ich do iných oblastí CNS. Vnútri uvedených systémov sa nachádzajú skupinky buniek funkčne delené podľa spojenia alebo mediátora na systémy/podsystémy: mozočkový systém a chemické systémy – cholinergický a monoaminergický (resp. katecholaminergický a seroninergický podsystém).
Retikulárna formácia plní mnohé aktivačné a inhibičné funkcie. Pôsobí budivo na mozgovú kôru a podieľa sa tak na kontrole cyklu bdenie – spánok, vytváraní stavu bdenia a je zapojená do prenosu pomalej nepresne lokalizovanej difúznej bolesti. Prerušenie RF vedie k strate vedomia.
Retikulárna formácia funguje ako „ strážnik“, ktorý dokáže zobudiť spiaci mozog v prípade zaznamenania dostatočne silného signálu z okolia. RF nepretržite registruje mozgovú aktivitu a je miestom, ktorým prechádzajú impulzy odosielané perifernými nervovými vláknami do CNS. Z dôležitých častí CNS sú privádzané do RF vzruchy, na ktoré bunky RF odpovedajú. Axóny buniek RF prebiehajú k vzdialenejším jadrám CNS a vydávajú aj početne rozvetvujúce sa kolaterály. Nervové vlákna tak vytvárajú povrazce, ktoré prechádzajú z jednej strany mozgového kmeňa na druhú, prebiehajú vzostupne alebo zostupne a spájajú jadrá havových nervov s mozgom, ako aj mozog s miechou a miechu s mozgom. Výrazné vnútorné prepojenie jadier RF tak zosilňuje podráždenie pôvodne jednoduchého nervového impulzu a vedie k aktivácií. RF je spojovacie a koordinačné ústredie motorických a senzitívnych nervov, zaisťuje mnohé prepojenia vnútri CNS. Je integračným centrom viscerálnych a svalových funkcií ( ovplyvňuje autonómne funkcie a riadi pohybovú aktivitu).
RF obsahuje nervové centrá dôležité pre ridanie vitálnych funkcií:
- centrum pre regulovanie dýchania (respiráciu)
- centrum pre vazomotoriku a činnosť srdca
- pneumotaktické centrum (zodpovedajú za pravidelnosť dýchania)
- centrum zvracania
RF je centrom dôležitých reflexov:
- obživných (prehĺtanie, slinenie,cicanie, sekrécia tráviacich štiav)
- obranných(žmurkanie, slzenie, kašlanie, zvracanie)
RF je zapojená ascendentne do riadenia senzitívnych a aktivačných systémov:
- vedie pomalú difúznu bolesť, udržiava stav bdenia a vedomie
RF je zapojená descendentne do riadenia motorických a autonómnych systémov:
- prináša motorické podnety z mozočka do nižších ústredí (miechy) a autonómne podnety z ústredia (podlôžka) na výkonný systém (sympatikus a parasympatikus).
Predný mozog a jeho vývin

Obrázok č. 5 Prosencephalon (Zdroj: Anatomy 3D Atlas)
Orel (2021) uvádza, že predný mozgový vačok prosencephalon sa rozdelí na diencephalon a telencephalon. Obe časti podliehajú ďalšiemu rastu, zmenám, vývinu a špecializácií. K ďalšiemu vývinu prosencephala pritom prispieva predovšetkým alárna a stropová platnička neurálnej trubice. Ku štruktúram, ktoré prislúchajú k diencephalu, radíme predovšetkým thalamus a hypothalamus, ale patria sem rovnako aj priľahlé oblasti epithalamus, metathalamus, subthalamus a očné vačky ako základ sietnice budúcich očí. Telencephalon sa následne sa vyvíja na dve mohutné mozgové hemisféry (pologuľe).
Medzimozog (diencephalon)
Prednú časť stropovej platničky z oblasti diencephala vytvoria spolu s vrstvou ependymových buniek strop III. mozgovej komory mozgovej. Z alárnej platničky medzimozgu vzniká thalamus a hypothalamus uložený nižšie:
- Párový thalamus – postupným rastom, zväčšovaním svojho objemu sa oba thalamy vyklenujú zo strán do III. mozgovej komory a zmenšujú ich veľkosť. V mieste kontaktu oboch thalamov vzniká u väčšiny mozgu rast – adhesio interthalamica. Thalamus sa stane hlavným prepojovacím ústredím mozgu. Postupne sa v ňom vyvinie rada špecializovaných thalamických jadier, ktorá bude prepojovaťspecifické informácie senzitívne, motorické a vegetatívne. Thalamy sa tak zapojujú do celej rady funkcií spojených s činnosťou zmyslov, riadením hybnosti, ale i do riadenia bdelosti, emócií, vegetatívnych reakcií a pod.
- Nepárový hypothalamus sa stane hlavným centrom riadenia vnútorných vegetatívnych alebo autonómnych funkcií, ktoré bude koordinovať a regulovať nezávisle našu vôľu. A teda postupne vznikne rada rôzne špecializovaných jadier, ktoré budú zasahovať do regulácie príjmu potravy a tekutín, tráviaceho pochodu, riadenia srdečnej činnosti, dýchania a telesnej teploty , rady ďalších telesných funkcií (vrátane sexuálnych) , ale i cirkardiánne rytmy (stredanie spánku a bdenia). Významne je zapojenie hypothalamu do správania a emočného prežívania, respektíve telesného prejavu emócií (nezabúdajme, že každá emócia má telesný doprovod, ktorý z veľkej časti sprostredkuváva práve hypothalamus). Niektoré neuróny hypothalamu budú nadané schopností neurokrinie (produkcia a transport chcemických látok hormonálnej povahy) a stanú sa producenty hormónov – konkrétne dvoch systémových hormónov (oxytocín a antidiuretický hormón/vazopresín) a rada regulačných hormónov (statín, liberín – viď. článok o oplodnení a vzniku embrya). Z výbežku diencephala smerujúceho dole sa vyvíja neurohypofýza – zadná časť podväzku mozgového. Nervovými vláknami sem budú z hypothalamu prúdiť obidva uvedené systémové hormóny (oxytocín a vazopresín). Neurohypofýza ich bude skladovať a postupne uvoľnovať do krvi. Pripomeňme všeobecne známy poznatok, že hypothalamus s spolu s hypofýzou vytvorí hypothalamu-hypofyzárny systém. Ten predstavuje najvyšší regulačný hormonálny systém, ktorého prostredníctvom bude mimo iného hormonálna produkcia rady iných žliaz s vnútornou sekréciou (ako je štítna žľaza, pohlavná žľaza, nadobličky). Dopady jeho činnosti sú veľmi široké.
Podľa Kluchovej (2015) je časť mozgu, ktorá sa rozprestiera okolo III. mozgovej komory. Štruktúry medzimozgu tvoria: lôžko, thalamus, madlôžko, epithalamus, zalôžko, metathalamus, podlôžko, hypothalamus a spodné lôžko, subthalamus. Malá brázda na laterálnej stene štrbinovitej III. komory – sulcus hypothalamicus – oddeľuje lôžko, nadlôžko a zalôžko dorzálne (hore) od podlôžka a spodného lôžka ventrálne (dole).
Lôžko (thalamus)
Najväčšia časť medzimozgu, diencephalon, tvorí lôžko, thalamus, ktoré tvoria dva ovoidné útvary zložené zo sivej hmoty. Útvary sú uložené po jednom na pravej a ľavej strane III. mozgovej komory.
Vonkajší opis
Lôžko, thalamus, predstavuje veľkú ovoidnú masu sivej hmoty, je to prevažne zhluk jadier, nuclei. Prístredná (mediálna) plocha lôžka je obrátená k III. mozgovej komore. Na jej hornom okraji je biely pruh vlákien – dreňový prúžok lôžka, stria medullaris thalami. Medzi dreňovými prúžkami oboch lôžok je rozopätá cievovková platnička, tela choroidea, ktorá zároveň tvorí strop III. mozgovej komory. Úpon tela choroidea na stria medullaris thalami sa nazýva prúžok lôžka, taenia thalami. Odtiaľ siaha zadná (dorzálna) , resp. horná plocha lôžka laterálne až po hraničný prúžok, stria terminalis, kde už susedí s ncl. caudatus. Dorzálna plocha lôžka je pozdĺžne rozdelená úponom plexus choroideus, resp. tela choroidea, na voľnú mediálnu časť a na laterálnu časť pokrytú platničkou, lamina affixa. Laterálne od lôžka je vnútorné púzdro, capsula interna, spredu (ventrálne), resp. zdola je lôžko oddelené od podlôžka prostredníctvom podlôžkovej brázdy, sulcus hypothalamicus. Užšia rostrálna časť lôžka tvorí predný hrbček lôžka, tuberculum anterius thalami. Zadná časť lôžka sa rozširuje do vankúša, pulvinar, ktorý zasahuje nad prístredné a bočné kolienkovité teleso, corpus geniculatum mediale et. laterale. Medzi lôžkom pravej a ľavej strany je často spojenie, medzilôžková hmota (priľnutie), adhesio interthalamica, bez bez funkčného významu.
Lôžko je zložené zo sivej hmoty – jadier a z bielej hmoty – vlákien. Biela hmota lôžka tvorí jeho vonkajšiu aj vnútornú vrstvu. Vonkajšia dreňová platňa, lamina medullaris externa, je biela vrtstva myelinizovaných vlákien na bočnej laterálnej ploche. (povrchu) lôžka, tesne pri capsula interna. Vnútorná dreňová platňa, lamina medullaris interna, je vertikálny pruh bielej hmoty, ktorý prechádza cez lôžko a smerom dopredu sa vidlicovito rozdvojuje, čím vytvorí vnútri každého lôžka útvar v tvare Y . Podľa jeho priebehu sa lôžko rozdeľuje na ventrálnu, mediálnu a laterálnu časť, ktoré obsahujú jednotlivé jadrá lôžka.
Vnútorná štruktúra – Sivá hmota lôžka
Anatomické delenie lôžka vyplýva z ich lokalizácie v závislosti od priebehu lamina medullaris interna.
Hlavné jadrá lôžka sú:
- ventrálne, ncl. anterior thalami
- mediálne, ncl. dorsomediálne, ncl. ventralis lateralis, ncl. ventralis posterolateralis, ncl. ventralis posteromedialis, ncl. corporis geniculati lateralis. Okrem týchto jadier sú mediálne uložené ncll. mediami, vnútri lamina medullaris interna sú uložené ncll. intralaminares a laterálne obkrúženie lôžka ncll. reticulares.
Funkčne sa jadrá lôžka na špecifické, nešpecifické a asociačné.
Špecifické jadrá
Ide o jadrá, ktoré sú spojené so špecifickými motorickými alebo senzitívnymi (senzorickými) oblasťami mozgovej kôry. Patria sem ventrálne jadrá – vpredu sú motorické jadrá ovplyvňujúce motorickú aktivitu, za nimi sú senzitívne jadrá prijímajúce senzitívne impulzy, celkom vzadu sú kolienkovité jadrá, ncll. geniculati. Špecifické jadrá ovplyvňujú jednak motorické funkcie mozgu ( motorické jadrá) a jednak jeho senzitívne funkcie (senzitívne jadrá). Súvis senzitívnych jadier so senzitívnymi funkciami je vyjadrený vedomým vnímaním prichádzajúcich z receptorov (dotyk, tlak, bolesť a pod.) Tu sa začína lôžkovokôrová dráha, smerujúca k príslušným jadrám v mozgovej kôre.
Stručné rozdelenie špecifických jadier
- Motorické jadrá: ncl. ventralis anterior dostáva aferentné vlákna z bazálnych ganglií a ncl. ventralis lateralis prijíma vlákna z mozočka. Eferentné vlákna z oboch jadier idú do motorickej kôry.
- Senzitívne jadrá: ncl. ventralis posterolateralis (aferenty z končatín a trupu) a ncl. ventralis posteromedialis (aferenty z tváre a hlavy). Obe jadrá sa uplatňujú pri vnímaní bolesti, vysielajú eferentné vlákna do primárnej senzitívnej kôry. K skupine senzitívnych jadier sa priraďujú aj ďalšie dve senzorické jadrá, ktoré sú uložené úplne vzadu, až pod pulvinar thalami. Sú to kolienkovité jadrá: ncl. corporis genuculati lateralis, s projekciou do primárnej zrakovej kôry. Obe jadrá sú považované už za súčasť zalôžka, metathalamus.
Nešpecifické jadrá
Ide o jadrá s nešpecifickým spojením s mozgovou kôrou (ncll. intralaminares, ncll. mediani et reticulares). Jadrá zaisťujú zameranie a koncentrovanie pozornosti na určité mentálne aktivity (napr. čítanie) a spolu s retikulárnou formáciou (RF) mozgového kmeňa sa podieľajú na udržiavaní bdelosti v dôsledku senzorickej aferentácie. Impulzy mozgového kmeňa sú prostredníctvom týchto jadier modulované, nadobúdajú aferentné rysy (príjemné alebo nepríjemné). Takto zmenené vnemy sa prepajajú cez limbickú kôru a zafixujú sa do pamäti. Uvedený modulačný efekt rovnako závisí od obojstrannej komunikácie s podlôžkom a limbickou kôrou.
- Ncll. intralaminares: sú lokalizované v bielej hmote lamina medullaris interna. Aferentné vlákna k nim prichádzajú z RF mozgového kmeňa, eferentné vlákna smerujú do bazálnych ganglií a difúzne do celej mozgovej kôry. Intralaminárne jadrá môžu byť považované za rostrálne pokračovanie RF z mozgového kmeňa a fungujú ako súčasť kôrového budivého mechanizmu (aferetného retikulárneho systému).
- Ncll. mediani: skupina buniek ležiaca na mediálnom povrchu lôžka, pozdĺž steny III. mozgovej komory. Funkčne sa im pripisuje úloha intergrácie viscerálnych aktivít. Tieto jadrá sú súčasťou ascendetného reitkulárneho systému.
- Ncll. reticulares: tvoria obal prednej časti lôžka, uložený na vonkajšej časti lamina medullaris externa, často prirovnávaný k vajcovej škrupinke. Má spojenia s inými jadrami lôžka, je akýmsi integrátorom jadier lôžka. Prostredníctvom ncll. reticulares reguluje mozgová kôra lôžkovú aktivitu, ale tieto jadrá ako jediné nemajú spätnú väzbu (teda nemajú vlákna smerujúce do mozgovej kôry).
Asociačné jadrá
Skupina asociačných jadier je obojstranne spojená aj s autonómnymi jadrami podlôžka, aj s limbickým systémom. Zároveň sú asociačné jadrá recipročne spojené s asociačnými oblasťami mozgovej kôry. Asociačné jadrá dodávajú senzitívnym vstupom emočné sfarbenie, napr. spojenie s kôrou čelového (frontálneho) citovo podfarbuje bolestivé podnety.
- Ncl. anterior:
- Ncl. dorsomedialis:
Súhrnne je funkčné rozdelenie jadier lôžka nasledovné:
Špecifické jadrá
- I. senzitívne – ncl. ventralis posteromedialis, ncl. ventralis posterolateralis, ncl. corporis geniculati medialis, ncl. corporis geniculati lateralis,
- II. motorické – ncl. ventralis anterior, ncl. ventralis lateralis
nešpecifické jadrá:
- asociačne jadrá: ncl. anterior thalami, ncl. dorsomadialis
Biela hmota lôžka
Medzi lôžkom a ostatnými časťami CNS existujú dôležité neurónové okruhy:
- Každé jadro lôžka (okrem retikulárneho) má recipročné spojenia s mozgovou kôrou, čím sa lôžko a kôra vzájomne ovplyvňujú
- Lôžko je dôležitou prepájacou stanicou pre dva senzitívno–motorické okruhy, do ktorých sú zapojené mozoček (1. okruh) a bazálne gangliá (2.okruh):
- 1. okruh: cerebello–rubro–thalamo–cortico–ponto–cerebellum
- 2.okruh: cortico–striato–pallido–thalamo–cortex
Spojenie medzi lôžkom a mozgovou kôrou je sprostredkované nervovými vláknami, ktoré vystupujú z laterálneho povrchu lôžka, prechádzajú cez bielu hmotu, capsula interna a rozbiehajú sa všetkými smermi do rôznych častí mozgovej kôry, pričom tvoria zväzky – lôžkové vetvenia, radiationes thalami:
- anteriores – spájajú lôžko s čelovou (frontálnou) kôrou mozgu
- centrales – smerujú z lôžka do temennej (parietálnej) mozgovej kôry
- posteriores – predstavujú spojenie senzorických jadier lôžka s kôrou mozgu: z corpus geniculatum mediale do spánkového (temporálneho) laloka a z corpus geniculatum laterale do záhlavového (okcipitálneho) laloka.
Funkcia lôžka
Komplexný pohľad na funkciu lôžka vyplýva z funkcií jeho jednotlivých jadier. Lôžko má celý rad funkcií (vrátane motorických), ale jeho najdôležitejšou funkciou je prijímanie senzitívnych (senzorických) informácií zo všetkých senzitívnych a senzorických systémov s jedinou výnimkou čuchového systému. Nervové vzruchy smerujú do mozgovej kôry, v nej sú prijímané do vedomia a analyzované. Každý takýto vzruch však musí prejsť cez thalamus, preto sa thalamus nazýva aj „bránou do vedomia“. Lôžko teda slúži ako prepájacia stanica vzruchov, ktorá „filtruje“ prichádzajúce informácie. Stáva sa tak posledným predstupňom mozgovej kôry. Informácie, ktoré lôžko neprepustí, nevnímame, teda akoby neexistovali (aj keď objektívne jestvujú). Touto filtráciou chránia jadrá lôžka mozgovú kôru pred nadmerným množstvom informácií. Lôžko takto výrazne ovplyvňuje reaktivitu mozgovej kôry. Eferentné dráhy lôžka smerujú prevažne do kôry mozgu, menšia časť ide do bazálnych ganglií a podlôžka.
Lôžko má významnú úlohu pri:
- integrácií senzitívnych a motorických systémov
- tvorbe osobnosti (prostredníctvom spojení ncl. dorsomedialis),
- úrovni vedomia (ncll. intralaminares ako pokračovanie formatio reticularis).
Stručne možno zhrnúť, že lôžko je ako celok súčasťou systému pre výber a ovplyvnenie informácií. Spolu s asociačnými oblasťami mozgovej kôry, s ktorými obojstranne komunikuje viacerými spojmi, sa významne podieľa na spracovaní aferentných impulzov na uvedomelé pocity a vnemy.
Nadlôžko – epithalamus
Dorzomediálna časť medzimozgu je tvorená nadlôžkom, epithalamus. Epithalamus sa nachádza pri zadnom okraji III. mozgovej komory, dorzomediálne od lôžka. Tvoria ho uzdička, habenula, uzdičkové jadrá, ncll. habenulares, spojka uzdičiek, commissura habenularum, zadná spojka, commissura posterior seu epithalamus, šuškovitá žľaza, glandula (gl.) pinealis. Endokrinná zložka nadlôžka je zapojená do limbického systému, tiež do mechanizmov ovplyvňujúcich spánok a cirkadiánne rytmy.
Zalôžko – metathalamus
Zalôžko, metathalamus, je uložené za lôžko, pod pulvinar thalami. V tejto časti medzimozgu sú uložené dva útvary: prístredné kolienkovité teleso, corpus geniculatum mediale, bočné kolienkovité teleso, corpus geniculatum laterale, v ktorých sa nachádzajú príslušné jadrá, nuclei. Ncl. corporis geniculati medialis je súčasťou sluchovej dráhy a ncl. corporis geniculati lateralis je zapojené do zrakovej dráhy.
Podlôžko – hypothalamus
– je najventrálnejšia uložená časť medzimozgu. Zúčastňuje sa na fungovaní troch systémov:
- autonómneho nervového systému
- endokrinného systému
- limbického systému
Vonkajší opis
Podlôžko je nepárová štruktúra, ktorá sa nachádza od obidvomi ovoidnými útvarmi lôžka, od ktorých je oddelené podlôžkovou brázdou, sulcus hypothalamicus. Tvorí spodinu III. mozgovej komory, laterálne čiastočne susedí so spodným lôžkom a čiastočne sa dotýka vlákien vnútorného púzdra, capsula interna. Rostrálnym ohraničením je hraničná platnička, lamina terminalis a na nej ležiaca predná spojka, commissura anterior. Kaudálne podlôžko prechádza do sivej hmoty stredného mozgu.
Na ventrálnej ploche podlôžka ( na spodine mozgu) sú uložené tieto štruktúry:
- skríženie zrakového nervu, chiasma opticum, tvorí spodinu zrakového zálivu, recessus opticus – je to záliv nad chiasma opticum do n. opticus
- lievik, infundibulum, z ktorého je na stopke zavesená podmozgová žľaza (hypofýza)
- popoĺavý hrboľ, tuber cinereum, medzi infundibulum a corpus mammillare
- bradavkovité teleso, corpus mammillare, a v ňom bradavkovité jadro, ncl. mammillaris
Vnútorná štruktúra – Sivá hmota podlôžka
Cez podlôžka prechádza oblúk, fornix. Je to dráha spájajúca hippocampus a corpus mammillare. Formix rozdeľuje podlôžko na mediálnu a laterálnu zónu. Obe zóny obsahujú viaceré jadrá.
Najdôležitejšie jadrá mediálnej zóny sú:
- ncl. preopticus reguluje vylučovanie gonadotropných hormónov
- ncl. suprachiasmaticus spojený so sietnicou kontroluje cirkadiánne rytmy (deň a noc), funguje ako nezávislé hodiny
- ncl. supraopticus a ncl. paraventricularis vylučujú hormóny – antidiuretický hormón (ADH) a oxytocín, ncl. paraventricularis vylučujú hormóny – antidiuretický hormón (ADH) a oxytocín, ncl. paraventricularis reguluje hladinu vody v tele, jeho poškodenie vedie k nadmernej spotrebe tekutín
- ncl. ventromedialis je centrum sýtosti, po podráždení utlmuje potrebu jedenia, poškodenie jadra, má za následok prejedanie (hyperfágiu) a obezitu
- ncl. dorsomedialis má úlohu pri prejavoch afektov a emócií, po jeho podraždení sa dostaví primitívne, grobianske správanie
- ncl. mammillaris je zapojený do limbického systému
- ncl. hypothalamicus anterior je zapojený do riadenia autonómnych parasympatikových funkcií, ako je termoregulácia a potenie, pri jeho poškodení dochádza k prehriatiu (hypertermii)
- ncl. hypothalamicus posterior riadi autonómne sympatikové funkcie, napr. rozširuje zrenice, zvyšuje krvný tlak, telesnú teplotu a svalové napätie prejavujúce sa trasom
- ncl. infundibularis a ncl. tuberalis sú zapojené dp funkcie adenohypofýzy, produkujú releasing faktory
Laterálna zóna podlôžka obsahuje jadrá uložené laterálne od fornix. Najvýznamnejším z nich je ncl. hxpothalamicus lateralis, centrum hladu, ktoré reguluje jedenie. Po jeho podráždení sa dostaví pocit hladu. Podlôžko má významnú úlohu pri regulácii zadnej časti podlôžka prevládajú sympatikové reakcie, zatiaľ čo stimulácia jeho prednej časti vyvolá parasympatikové reakcie, zatiaľ čo stimulácia jeho prednej časti vyvolá parasympatikové reakcie. Činnosť podlôžka pri zabezpečovaní sexuálnych funkcií sa uskutočňuje jeho intergračným pôsobením (ako na križovatke) CNS a hormonálnej sekrécie. Zodpovednosť za vznik ženského sexuálneho správania sa prisudzuje zadnému podlôžku, zatiaľ čo mužské sexuálne správanie je ovplyvňované preoptickou oblasťou.
Biela hmota podlôžka
Do podlôžka prichádzajú aferentné vlákna:
- viscerálne a somatické (cez retikulárnu formáciu)
- z pokrývky (tegmentum) stredného mozgu
- z hipokampu (cez fornix)
- z amygdaly (cez stria terminalis a vemtrálnu amygdalofugálnu dráhu)
- cez septa (cez facsiculus telencephalicus medialis)
- z čuchovej kôry
- z lôžka (z ncl. dosrdomedialis)
- z čelového laloka mozgovej kôry (priamo do podlôžka)
Hlavné eferentné vlákna z podlôžka smerujú:
- do lôžka, cez tractus mammillothalamicus
- do stredného mozgu, cez tractus mammillotegmentalis
- k jadrám mozgového kmeňa a miechy, cez fasciculus longitudinalis posterior (nazývaný aj fasciculus longitudinalis posterior (nazývaný aj fasciculus longitudinalis dorsalis)
Funkcia podlôžka
Podlôžko spája a zjednocuje descendentné signály z mozgovej kôry (prostredníctvom amygdaly a hipokampovej formácie) a ascendentnými signálmi z miechy a mozgového kmeňa. S takto upravenou informáciou potom podlôžko riadi viscerálne funkcie v súvislosti so správaním, ktoré zaručuje prežitie jedinca a druhu, napr. koordinácia sexuálneho správania s neuroendokrinnou reguláciou reprodukčným orgánov zaisťuje prežitie druhu. Koordinácia pitia a jedenia s gastrointestinálnymi funkciami zaisťuje prežitie jedinca. Podobne termoregulačné deje musia byť koordinované s teleným metabolizmom, ako aj je periférne napätie ciev a potenie spojené s telesnou teplotou. Podlôžko a limbický systém taktiež ovplyvňujú emocionálne správanie – hnev, zúrivosť, pokojnosť, strach a spoločenskú príťažlivosť. Pri poškodení prednej časti lôžka dochádza k prevahe funkcií jeho zadnej časti, následkom čoho sa u jedinca prejaví násilie a agresivita (škodoradosť,zlomyselnosť, ničomnosť, nemravosť) a to aj voči milovaným osobám. Kontrola uvedených komplexných dejov si vyžaduje súčinnosť endokrinného systému a autonómneho nervového systému (ANS). Podlôžko teda funguje ako nadradené centrum endokrinnej aktivity a aktivity autonómneho nervového systému:
- reguluje endokrinnú aktivitu sekréciou neuroendokrinných produktov do krvi
- kontroluje ANS nervovými dráhami cez formatio reticularis a tiež priamo nervovými dráhami k parasympatikovým a sympatikovým jadrám hlavových nervov a miechy cez fasciculus longitudinalis posterior a cez zostupné dráhy retikulárnej formácie). Týmto pôsobením podlôžko následne významne ovplyvňuje aj aktivitu limbického systému.
Podlôžko a emócie
Podlôžko reguluje autonómne prejavy nervových impulzov, ktoré vyvolávajú fyzické prejavy emócií: zrýchlenie srdcovej činnosti, zvýšenie tlaku krvi, začervenanie alebo zblednutie tváre, potenie, husia koža, suchosť v ústach a gastrointestinálne poruchy. Mozgová kôra je zodpovedná za vznik subjektívnych pocitov sprevádzajúcich emočné zážitky. Okrem toho mentálne procesy v mozgovej kôre, ktoré majú silnú emocionálnu zložku, môžu vyvolať ďalšie reakcie podlôžka. Nervové dráhy, ktoré spájajú mozgovú kôru a podlôžko, sú zapojené do emočných reakcií. Pri týchto reakciách zohráva významnú úlohu hipokampová formácia. štruktúry súvisiace s čuchovým systémom a ďalšie štruktúry limbického systému. Zapojenie podlôžka do emočných procesov sa uskutočňuje aj súhrou s ďalšími šturktúrami mozgu. Na konci týchto procesov sa dostavia somatomotorické reakcie a reakcie autonómneho nervového systému, teda vegetatívne reakcie. Vegetatívne reakcie podlôžka sú výrazným prejavom organizmu na jeho psychickú aj emocionálnu záťaž.
- Psychická záťaž organizmu vyvoava rad vegetatívnych reakcií, ktoré majú podobný priebeh ako pri fyzických reakciách, napr.: zvýšené vylučovanie adrenalínu v krvi, zvýšená srdcová činnosť, zrýchlenie dýchania, zvýšené prekrvenie kože spolu so zvýšeným vylučovaním potu
- Emocionálna záťaž (napr. strach, zlosť, hnev) má za následok rozvoj podobných reakcií ako pri psychickej záťaži
- Pri pocite strachu, vzrušenia a počas život ohrozujúcej situácie je v podlôžku výrazne stimulivaný sympatikový nervový systém sprevádzaný prejavmi tachykardie, hyperventiláciou, potením a pod.
Spodné lôžko
Laterálne od podlôžka je uložené spodné lôžko, subthalamicus. Patrí k nemu jadro spodného lôžka, ncl. subthalamicus, ktoré je uložené pod lôžkom a ventrálne. Medzi ním a lôžkom je neurčité pásmo, zona incerta, ktoré tvorí mediálne pokračovanie ncll. reticulares. Cez spodné lôžko prechádza niekoľko zväzkov vlákien spájajúcich bazálne gangliá a jadrá lôžka.
Koncový mozog (telencephalon) ako súčasť predného mozgu
Telencephalon je posledným oddielom mozgového vačku. Stane sa vôbec najmohutnejšou časťou ľudského mozgu. Práve štruktúry koncového mozgu najviac odlišujú mozog človeka od mozgu iných cicavcov a umožňujú realizovať špecificky ľudské funkcie mozgu a psychiky. Už začiatkom piateho týždňa vnútromaternicového života vznikajú základy mozgových pologúľ (mozgových hemisfér) ako vyklenutie vonkajších strán prosencephala. Už na začiatku nájdeme v ich vnútri primitívne dutiny, ktoré predstavujú základ dvoch postranných dutín (ventruli laterales). Zhruba v polovici druhého mesiaca bazálnej časti oboch hemisfér mohutnie a ako gangliové hrboľky sa vykleňujú do priestoru primitívnych postranných komôr. Z tejto časti sa postupne vyvinú mohutné podkôrové útvary koncového mozgu – bazálne gangliá/jadrá. K bazálnym gangliám radíme niekoľko štruktúr uložených v hĺbke mozgových hemisfér. Sú prepojené vzájomne i s oblasťami motorickej kôry a thalamov. Hrajú veľmi významnú rolu v riadení motoriky, ale plnia rovnako asociačné a pamäťové funkcie. Po novom sa im priraďuje tiež určitý podiel v pôvode pozornosti a môžu zrejme zohrávať tiež určitú rolu v utváraní osobnosti (pokiaľ si uvedomíme, ako zložitá a komplexná osobnosť človeka je). Zatiaľ čo vonkajšia laterálna strana mozgových hemisfér mohutnie a rastie , vnútorná mediálna stena zostáva tenká, pretože tu nedochádza k množeniu neuroblastov. Je de facto tvorená len vrstvou ependýmových buniek. Tie spolu s vaskularizovaným tkanivom vytvorí choroideálny plexus postrannej komory (plexus choroideus ventriculi lateralis). Keďže sa hemisféry intenzívne zväčšujú a mohutnejú, pomaly sa usadia aj na medzimozog a dostávajú sa do priameho kontaktu s oboma thalamami. Ich rast sa nezastavuje a tak prerastajú i stredný mozog a základ mozočka. Mozgové hemisféry rastú smerom dopredu, dozadu a dole, čo vedie ku vzniku mozgových lalokov – čelný, spánkový, záhlavový, temenný. Oblasť koncového mozgu, ktorá susedí so žíhaným telesom, sa vyvíja o niečo pomalšie ako ostatné časti, ktoré ju začnú čoskoro prerušovať. Nazýva sa inzula (ostrov) a ku koncu tehotenstva bude prekrytá čelovým, spánkovým a temenným lalokom. Zostane tak skrytá ako ostrovný lalok (lobus insularis), táto časť mozgu bude hrať rolu v integrácií regulácií, riadení vnútorných funkcií a bude zasahovať tiež do prežívania a emócií.
Známy prehľad 5 mozgových lalokov, ktoré sa vyvíjajú v priebehu vnútromaternicového života:
- lalok frontálny/čelový (lobus frontalis)
- lalok temenný/parietálny (lobus parietalis)
- lalok spánkový/temporálny (lobus temporalis)
- lalok okcipitálny (lobus occipitalis)
- lalok ostrovný/inzulárny (lobus insularis)
Najprv je povrch mozgových hemisfér a teda ich jednotlivých lalokov hladký. Akokoľvek sa neuroblasty pri povrchu mozgových hemisfér intenzívne zmnožia, zväčšuje sa postupne povrch mozgovej kôry. To vedie, ku vzniku vyklenutých mozgových závitov – gyrov, ktoré sú od seba oddelené ryhami a hlbokými brázdami. Proces členenie mozgových pologúľ do závitov oddelených ryhami sa nazýva gyrifikácia. Centrálna ryha (sulcus centralis) oddeľuje čelný a temenný lalok a bočná ryha (sulcus lateralis) položená medzi čelným a spánkovým lalokom sa objavuje už koncom šiesteho týždňa. Proces gyrifikácie postupuje v priebehu vnútromaternicového života ďalej – mozog novorodenca má základnú gyrifikáciu v podstate na úrovni odpovedajúceho mozgu dospelého človeka.
Masa koncového mozgu je tvorená šedou a bielou mozgovou hmotou:
- Šedá hmota mozgová – predstavuje zhromaždenie tiel nervových buniek, vzniká zmnožením, migráciou a diferenciáciou neuroblastov.
- Tvorí povrchovú vrstvu mozgových hemisfér v podobe mozgového plášťa (pallium), ktorý je základom mozgovej kôry (cortex cerebri) a leží v hĺbke koncového mozgu v podobe podkôrových (subkortikálnych) útvarov, respektíve bazálnych ganglií (jadier). Na reze mozgu je celkom dobre viditeľná sivastá farba.
- Biela hmota mozgová – je tvorená predovšetkým nervovými vláknami. Nájdeme ich v hĺbke mozgových hemisfér, pod mozgovou kôrou. Na reze mozgu sú biele farby. Rozvíja sa najmä rastom výbežkov a ich myelinizáciou, teda vytvorením obalov.
Mozgový plášť (pallium) sa člení na tri časti, z ktorých vznikne allocortex, mesocortex a neocortex – tri typy mozgové kôry, ktoré sa líšia počtom bunečných vrstiev a zložitostí svojho usporiadania:
Allocortex – odhadom necelých 5 % celkovej plochy mozgovej kôry. Tvorí čuchovú, kôrovú oblasť a kôrovú časť limbického systému. Obsahuje tri vrstvy neurónov.
Mesocortex – predstavuje prechodový lem medzi trojvrstvovým allokortexom a šesťvrstvovým neokortexom, obsahuje teda štyri alebo päť vrstiev neurónov.
Neocortex – reprezentuje jednoznačne dominantnú časť mozgovej kôry (tvorí viac ako 90%). Ide o najmohutnejšiu a vývinovo najmladšiu kôru, ktorá je tvorená celkom šiestimi vrstvami neurónov.
Rozdielne oblasti mozgovej kôry obsahujú jednak rozdielny počet vrstiev neurónov, podľa toho, či ide o allocortex, mesocortex alebo neocortex, ale tiež nervové bunky odlišnej veľkosti, tvaru, stavby a funkcie. Početnosťou však prevažujú dva typy – pyramídové a granulárne.
Pyramídové bunky – dostali meno podľa ich skutočného tvaru pyramídy. Jedná sa o nervové bunky excitačné, ktoré fungujú ako neuróny projekčné (vysielané axóny do podkôrových štruktúr mozgu), neuróny asociačné (ktoré prepájajú rôzne oblasti mozgovej kôry v jednej mozgovej poľguľe) a neuróny komisurálne (vysielajú výbežky do protiľahlej hemisféry). Pyramídové neuróny sa významne uplatňujú v motorických dráhach, ktoré sú určené k riadeniu voľnej hybnosti.
Granulárne (zrnité) bunky – majú menšie telo a v mikroskope pripomínajú drobné zrniečka (granule). Fungujú predovšetkým ako interneuróny so spájajúcou funkcií. Ich axón je krátky a neopúšťa oblasť mozgovej kôry. Granulárne neuróny môžu byť ako excitačné , tak inhibičné. Sú významne zapojené predovšetkým do spracovania senzitívnych informácií.
Funkcie jednotlivých neurónov a ich synaptické kontakty sú základom komplexnej mozgovej činnosti. Vedľa spojení jednotlivých neurónov na mikroskopické úrovni sa v mozgu postupne vytvárajú významne dráhy, ktoré navzájom prepájajú obe mozgové poľogule – mozgové komisúry (commisurae cerebri). Najväčšie sú tri z nich, ktoré dosahujú veľkosť, že sú veľmi dobre viditeľné aj ľudským okom:
Corpus callosum – kalózne teleso, je najmohutnejšou, najvýznamnejšou komisúrou mozgu – preto ju uvádzame na prvom mieste, z časového hľadiska vzniká až ako posledná. Vyvíja sa od desiateho týždňa vnútromaternicového života a postupne a zväčšuje a rozširuje. Obrovským množstvom nervových vlákien (v rade stoviek miliónov) navzájom prepája rozsiahle oblasti pravé a ľavé mozgové hemisféry.
Commisura anterior – predná komisúra, vyvíja sa z komisurálnej platničky, čo je mohutnejšia lamina terminalis, ktorá sa objavuje v piatom týždni vnútromaternicového života. Táto komisúra prejavuje najmä kôrové oblasti čuchu a tiež laloky spánkové.
Commisura fornicis – vzniká vnútromaternicovo ako druhá v poradí. Prepája navzájom štruktúry limbického systéma, konkrétne pravý a ľavý parahipokampálny závit (gyrus parahippocampalis) a oblasť pravého a ľavého hipokampu (hippocampus).
Už na konci tehotenstva je nervový systém a sústava zmyslov na takej úrovni vývinu, že sú hotové vykonávať svoje funkcie. Kognitívne, zmyslové, pamäťové a emočné schopnosti sú pred narodením na omnoho vyššej úrovni, ako sa skôr predpokladalo. Už na konci vnútromaternicového života, teda ešte pred narodením, môže plod vnímať radu podnetov a učiť sa . Bezpečne rozpozná matkin hlas, vníma činnosť matkino srdca a ostatných orgánov. Registruje veľa zvukov , rovnako aj taktilné podnety z vonka a reaguje na ne. Mnohé emočné a telesné stavy matky sa môžu prenášať na plod, ktorý je vníma a reaguje na ne. Dieťa sa tak nepochybne rodí s určitým bazálnym základom a výbavou ovplyvnenou genetickým základom. Teda novorodenca nemôžme považovať za „nepopísaný čistý list/tabula rasa.“
Ako uvádza Boleková (2015) koncový mozog, telencephalon, tvorí časť predného mozgu a je najväčšou časťou mozgu. Je zložený z dvoch poľogúl, hemisphareiae cerebri. Na povrchu koncového mozgu je vrstva sivej hmoty, ktorá tvorí mozgovú kôru, cortex cerebri, rozbrázdenú do charakteristických mozgových závitov. Vnútri koncového mozgu sa nachádza biela hmota, corpus medullare, v ktorej sú uložené zhluky sivej hmoty, bazálné gangliá, nuclei basales. Súčasťou koncového mozgu je aj čuchový mozog, rhinencephalon, a časť limbického systému, obe časti navzájom spolu úzko súvisia.
Všeobecná charakteristika
– je vývinovo najmladšou časťou mozgu. Je tvorený mozgovými pologuľami (hemisférami), hemispharae cerebri, ktoré pokrýva povrchová vrstva sivej hmoty, mozgová kôra, cortex cerebri. Vnútri hemisfér sa rozprestiera biela hmota, dreňové teleso, corpus medullare, ktorá je oveľa hrubšia ako mozgová kôra. Biela hmota je zložená z vlákien, ktoré sú súčasťou nervových dráh. V podkôrových častiach hemisfér sa v bielej hmote nachádza vnútorná sivá hmota. Tvorí veľké jadrá, nazývané bazálné gangliá, ncll. basales, ktoré sú zložené z tiel nervových buniek. Jedna hemisféra má u človeka vždy dominantné postavenie a nazýva sa dominantná hemisféra. U pravákov je to ľavá hemisféra, ktorá riadi reč, písanie a počítanie, kým pravá hemisféry emočné, neverbálne myslenie a umelecké schopnosti, u ľavákov je to naopak. Nedominantá hemisféra sa naviac uplatňuje pri ukladaní trvalých pamäťových stôp. Najstaršou časťou koncového mozgu je čuchový mozog, rhinencephalon, ktorý je úzko funkčne úzko spojený s limbickým systémom mozgu, systema limbicum, centrom emócií, správania sa a pamäti. Vnútorné štruktúry hemisfér prechádzajú do medzimozgu a cez pedunculi cerebri sa spájajú s mozgovým kmeňom. Koncový mozog je rozdelený hlbokou pozdĺžnou mozgovou štrbinou, fissura longitudinalis cerebri, na pravú a ľavú hemisféru. Vpredu sa na každej hemisfére nachádza predný, čelový pól, polus anterior seu frontalis, vzadu zadný, záhlavový pól, polus posterir seu occipitalis. Spánkový pól, polus temporalis, je na prednej časti spánkového laloka. Každá hemisféra má tri plochy: komplexnú hrnobočnú plochu, facies superolateralis, plochú prístrednú plochu, facies medialis, a dolnú plochu, facies inferior, ktoré navzájom oddeľujú tri okraje: horný okraj, margo superior – medzi facies superolateralis a facies medialis, dolný prístredný okraj, margo inferomedialis – medzi medialis a facies inferior a dolný bočný okraj, margo nferomedialis – medzi facies medialis a facies inferior a dolný, bočný okraj, margo inferolateralis – medzi facies superolateralis a facies inferior. Obe hemisféry navzájom spája svorové teleso, corpus callosum, ktoré je výraznou štruktúrou na facies medialis . Pod corpus callosum sa nachádza oblúk, fornix, ktorý čiastočne sleduje zakrivenie svorového telesa. Jeho predný úsek, nazývaný stĺpec oblúka, columna fornicis, je súčasťou III. mozgovej komory a tvorí predné ohraničenie medzikomorového otvoru, foramen interventriculare. Oblúk pokračuje nahor telom oblúka, corpus fornicis, opušťa prístrednú plochu hemisféry a pokračuje ramenom oblúka, crus fornicis, dozadu a nadol do dolného rohu bočnej mozgovej komory. Medzi prednými časťami oblúka a svorového telesa sa rozprestiera platňa priezračnej priehradky, lamina septi pellucidi, pravá a ľavá lamina ohraničujú priezračnej priehradky, cavum septi pellucidi.
Mozgová kôra – cortex cerebri
– tvorí povrchovú časť koncového mozgu a u človeka je usporiadaná do viacerých vrstiev, v závislosti od stupňa vývinu príslušnej oblasti mozgu.
Funkčné kôrové oblasti
Pre kompletnú verziu cytoarchitektonickej mapy mozgovej kôry dospelého človeka opísal začiatkom 20. storočia (1909) nemecký neurológ Korbinian Brodmann. Mozgovú kôru rozdelil na 11. oblastí, regiones a 52 menších morfologicky odlišných políčok, areae. Táto mapa bola viackrát modifikovaná, ale jej podoba sa zachovala a používa sa dodnes. Nové metódy klinického výskumu prinášajú stále novšie informácie o type nervových a podporných buniek a ich funkcií, skúmajú synaptické spojenia a vlásočnice. Vytvárajú sa tak nové cytoarchitektonické, glioarchitektonické, myeloarchitektonické a angioarchitektonické mapy. Na základe podložených klinických a experimentálnych poznatkov predstavuje cortex cerebri unikátne sídlo ľudského procesu poznávania a myslenia. V mozgovej kôre sú lokalizované väčšie a menšie funkčné oblasti, do ktorých vstupujú informácie z receptorov. Podrobnou analýzou a následnou syntézou priatej informácie sa formuje vhodná a cielená odpoveď, reakcia. Okrem senzitívnych kôrových oblastí sú v kôre uložené aj motorické oblasti, ktoré riadia vôľové, vedomé pohyby a vedomú činnosť. Rozsiahlejšia časť kôry patrí asociačným oblastiam, ktoré umožňujú prepojenie rôznych častí mozgovej kôry a koordinácia senzitívnej funkcie a motorickej činnosti.
Najdôležitejšie funkčné kôrové oblasti
Motorické oblasti: lokalizované pred sulcus centralis
Primárna motorická oblasť – M1, area motoria, je lokalizovaná v gyrus precentralis, dráždením oblasti dochádza ku kontrakcii priečne pruhovaných svalov a pri jej poškodení dochádza k ochrnutiu (paralýze) priečne pruhovaných svalov na opačnej strane tela.
Sekundárna motorická oblasť – M2, je v oblasti gyrus frontalis superior, pripravuje a iniciuje pohyb.
- Premotorická oblasť – PM, area premotoria, zahŕňa predný úsek gyrus precentralis a zadné úseky gyri frontales, ovplyvňuje prípravu a zmenu pohybu.
- Frontálne okohybné pole – FEF , je uložené pred premotorickou oblasťou, v gyrus frontalis medius, zodpovedá za koordináciu pohybu očí, hlavy a krku.
- Prefrontálna oblasť – sa nachádza pred premotorickou oblasťou, má spojenia a lôžkom a podlôžkom, ovplyvňuje vnímanie, schopnosť koncentrácie a učenia, zodpovedá za plánovanie, vôľové konanie a viaceré aspekty integrity jedinca.
- Motorická rečová oblasť – je lokalizovaná v pars triangularis gyri frontalis inferioris, resp. v operculum frontale, zodpovedá za správnu formuláciu slov synchronizovaním svalov riadacich dýchanie a tvorbu hlasu (fonáciu).
Senzitívne (senzorické) oblasti – lokalizované za sulcus centralis
- Primárna senzitívna oblasť – S1, area sensoria, zaberá gyrus potcentralis a lobulus paracentralis, je zodpovedná za uvedomovanie si senzitívnych pocitov z tela.
- Sekundárna senzitívna oblasť – S2, oblasť okolo hornej časti sulcus lateralis, reaguje na podnety kožnej citlivosti.
- Senzitívna rečová oblasť (Wernickeho senzitívne centrum reči) – je lokalizovaná v zadnom úseku gyrus temporalis superior a v priľahlej temennej oblasti, zodpovedá za porozumenie hovorenej reči a ovplyvňuje tak proces učenia, väčšina prichádzajúcich senzitívnych podnetov je trasformovaná podoby, u pravákov je lepšie vyvinutá v dominantnej, ľavej hemisfére.
- Primárna sluchová oblasť– A1, area acustica, zaberá gyri temporales transversi, prijíma vlákna z corpus geniculatum mediale, ktoré formujú radiato acustica.
- Primárna zraková oblasť – V1, area visualis, je oblasť okolo sulcus calcarinus, zasahuje do oblasti sulcus lunatus (tvorí jej ohraničenie) a do gyrus lingualis, prijíma vlákna z corpus geniculatum laterale, ktoré formujú radiatio optica.
- Zraková psychická oblasť, area parastriata, obklopuje primárnu zrakovú oblasť a je odpovedná za vznik vizuálnych dojmov, ktoré sú dôležité pre poznanie a identifikáciu.
- Chuťová oblasť, area olfactoria, leží na dolnej ploche lobus temporalis v oblasti rostrálneho a laterálneho úseku, uncus gyri parahippocanpalis.
Biela hmota – corpus medullare
Koncového mozgu vypĺňa priestor medzi mozgovou kôrou a jadrami koncového mozgu. Polooválne centrum, centrum semiovale, spája s druhostranným centrom svorové teleso, corpus callosum. Zo svorového telesa prebiehajú vlákna vejárovito a tvoria lúčovito vetvenie svorového telesa, radiatio corporis callosi. Bielu hmotu tvorí, okrem zastúpenia gliových buniek a tenkostenných kapilár, veľký počet myelinizovaných výbežkov neurónov. Jenotlivé vlákna lokalizované v corpus medullare tvoria dráhy asociačné, komisurálne a projekčné.
Biela hmota koncového mozgu, corpus medullare, vypĺňa priestor medzi mozgovou kôrou a jadrami koncového mozgu (bazálne gangliá). Na horizontálnom reze mozgom tesne nad corpus callosum pripomína biela hmota tvarom poloovál s nazýva sa centrum semiovale. Zo svorového telesa prebiehajú vlákna vejárovito do kôry a tvoria lúčovité vetvenie svorového telesa, radiatio corporis callosi.
Bielu hmotu tvorí, okrem zastúpenia gliových buniek a tenkostenných kapilár, veľký počet myelinizovaných výbežkov neurónov, ktoré vytvárajú:
- asociačné dráhy, prebiehajúce medzi odlišnými časťami tej istej hemisféry a tvoriace zväzky: opasok, cingulum, hákovitý zväzok, fasciculus uncinatus, oblúkovitý zväzok, fasciculus arcuatus, horný a dolný pozdĺžny zväzok, fasciculus longitudinalis superior et inferior
- komisurálne dráhy, spájajúce obidve hemisféry navzájom, svorové teleso, corpus callosum, predná spojka, commissura anterior, zadná spojka, commissura posterior, svojka uzdičiek, commissura habenularum a spojka oblúka, commissura fornicis
- projekčné dráhy, prichádzajúce z nižších oblastí CNS k neurónom vyšších oblastí CNS – do kôry koncového mozgu a tiež zostupujúce z vyšších oblastí CNS k neurónom nižších oblastí CNS
Na horizontálnom reze v úrovni genu et splenium corporis callosi vlákna oboch častí formujú oblúkovité štruktúry otvorené dopredu, tzv. malé kliešte, forceps minor seu fontalis, a otvorené dozadu, tzv. veľké kliešte, forceps major seu occipitalis. Corpus callosum pokrýva tenká vrstvička sivej hmoty, sivá zásterka, indusium griseum, ktorá spája gyrus paraterminalis a gyrus dentatus postredníctvom povrchovo uložených párových prístredných a bočných pozdĺžnych prúžkov bielej hmoty, striae longitudinales mediales et laterales. Tenká vrstvička bielej hmoty, alveus hippocampi, vlákna ktorej sa zbiehajú mediálne do strapca, fissura hippocampi, pokrýva útvar lokalizovaný v dolnom rohu bočnej mozgovej komory, nazývaný hippocampus.
Podľa priebehu k podkôrovým centrám alebo do nižších centier na úrovni mozgového kmeňa a miechy projekčného dráhy obsahujú obojsmerne:
- krátke projekčné vlákna (spájajúce kôru mozgu s bazálnymi gangliami, lôžkom a podlôžkom),
- dlhé projekčné vlákna (spájajú kôru mozgu s mozgovým kmeňom a miechou).
Terminálne úseky projekčných dráh spájajú podkôrové a kôrové centrá medzimozgu a koncového mozgu a tvoria vajcovitý komplex vlákien, lôžkovokôrové vetvenie, radiatio thalamocorticalis. Vlákna spájajú aj kôrové a podkôrové centrá koncového mozgu a medzimozgu a vytvárajú vetvenie prúžkovaného telesa, radiatio corporis striati.
Okolo jadier vnútri každej hemisféry tvorí biela hmota puzdrá, capsulae. Najväčšie vnútorné púzdro, capsula interna, je tvorená vláknami dlhých projekčných dráh, ktoré v ňom prebiehajú, topicky usporiadané. Vnútorné púzdro, capsula interna, má na horizontálnom reze hemisférou tvar širokého písmena V, otvoreného smerom laterálne, od ktorého sa rozostupujú nervovú vlákna smerujú ku kôre, tvoriac vejárovitý útvar, lúčovitý veniec, corona radiata. Vnútorné púzdro tvoria tri časti:
- predné rameno vnútorného púzdra, crus anterior caplsulae internae, lokalizované medzi hlavou chvostnatého jadra, caput ncl. caudati a šošovkovitým jadrom, ncl. lentiformis:
- koleno vnútorného púzdra, genu caplsulae internae
- zadné rameno vnútorného púzdra, crus posterius capsulae internae, loalizované medzi thalamus a ncl. lentiformis
Laterálne od ncl. lentiformis sa nachádza vonkajšie púzdro, capsula externa, ktoré tvorí biela hmota medzi ncl. lentiformis a závorou, claustrum. Najvzdialenejšie púzdro, capsula extrema, je uložené laterálne od claustrum a tvorí okrajové ohraničené bielej hmoty. Cez capsula interna prebiehajú dôležité aferentné a eferentné vlákna nervové dráhy usporiadané v nasledovnom poradí:
cez crus anterius capsulae internae:
- čelovomostová dráha , tractus frontopontinus, súčasť tractus contico-ponto-cerebellaris:
- predný lôžkovokôrový zväzok, fasciculus thalamocorticalis anterior (radiatio thalamus anterior)
cez genu capsulae internae:
- kôrovojadrová dráha , dráha corticonucleris, smerujúca k motorickým jadrám hlavových nervov (pre svaly hlavy a krku)
cez predný úsek crus posterius capsulae internae
kôrovomiechová dráha, tractus corticospinalis, smerujúca k motorickým jadrám miechy (pre svaly hornej končatiny a trupu)
cez centrálny úsek crus posterius capsulae internae:
- kôrovočervená dráha, tractus corticorubralis,
- kôrovosieťková dráha, tractus corticoreticularis
- kôrovomiechová dráha, tractus corticospinalis
- ústredný lôžkovokôrový zväzok, fasciculus thalamocorticalis centralis (radiatio thalami centralis), v oblasti ktorého prebieha aj kôrovolôžková dráha, tractus cortiicothalamicus, obe dráhy tak tvoria obojsmerné zoskupenie nervových vlákien
cez zadný úsek crus posterius capsulae internae v pars retrolentiformis
- zadný lôžkovokôrový zväzok, fasciculus thalamicocorticalis posterior (radiatio thalami posterior), temenno-záhlavno-mostová dráha, tractus-occipito-pontinus
v zadnom úseku crus capsulae ibternae:
- spánkovomostová dráha, tractus temporopontinus
- koncové úseky, senzorických dráh (sluchovej a zrakovej), sluchové vetvenie, radiatio acustica, rakové vetvenie, radiatio optica
Fasciculus thalamicocorticalis anterior et posterior predstavujú koncové úseky senzitívnych dráh.
Bazálne gangliá – nuclei basales
Termínom bazálne gangliá označujeme zhluk jadier uložený v hĺbke mozgu v bielej hmote, laterálne od medzimozog. Funkčne sú tieto jadrá zapojené do iniciácie a kontroly vôľových pohybov. Medzi bazálne gangliá patria štruktúry koncového mozgu – corpus striatum medzimozgu – ncl. subthalamicus a stredného mozgu – substantia nigra. Chvostnané jadro, ncl. caudatus a šošovkovité jadro, ncl. lentiformis, patria k najväčším celkom tvoriacim bazálne gangliá. Bazálne gangliá predstavujú zhluk rôzne tvarovaných jadier, uložený v bielej hmote mozgu. Vytvárajú ich štruktúry koncového mozgu, medzimozgu a stredného mozgu.
Prúžkované teleso – corpus striatum
Prúžkované teleso, corpus striatum, je zložené, z ncl. caudatus a ncl. lentifirmis. Tieto dve veľké hmoty jadier sú uložené vnútri mozgových hemisfér, medzi lôžkom a mozgovou kôrou (neocortex).
Chvostnaté jadro – nucleus caudatus
Ncl. caudatus má podobu zatočenej masy sivej hmoty, uloženej v stene bočnej mozgovej komory. Rozdeľuje sa na tri časti: hlava, caput, telo, corpus a chvost, cauda. Najväčšiu časť tvorí caput, ktoré vyčnieva do predného rohu bočnej komory. Smeruje dozadu, kde sa zužuje a na úrovni medzikomorového otvoru, foramen interventriculare, prechádza do corpus. V ďalšom priebehu sa stáča nadol a prechádza do spánkového laloka ako cauda, kde sa končí spojením s mandľovitým telesom, corpus amygdaloideum.
Šošovkotivé jadro – nucleus lentiformis
- Ncl. lentiformis sa tvarom podobá na šošovicu alebo lentilku, tvorí ho škrupina, putamen a bledá guľa, globus pallidus. Putamen sa nachádza laterálne, je uložený medzi capsula externa a globus pallidus. Lokalizácia globus pallidus je potom medzi putamen a capsula interna. Ncl. lentiformis je od lôžka oddelený bielou hmotou capsula interna prostredníctvom zadného ramena, crus posterius capsulae internae, a od ncl. caudatus prostredníctvom predného ramena, crus anterius capsulae internae.
- Ncl. caudatus a putamen spolu morfologicky, funkčne a z vývinového hľadiska, klinicky sú nazývané striatum , neostriatum. Naproti tomu, vývinovo starší globus pallidus, nazývaný aj ako pallidum, je štruktúra odlišná morfologicky aj fyziologicky. Vplyv neurónov globus pallidus na cieľové štruktúry je inhibičný.
Jadro spodného lôžka – nucleus subthalamicus
Ncl. subthalamicus je najväčšie jadro spodného lôžka – štruktúry klinovitého tvaru, uloženej ventrálne od lôžka a laterálne od podlôžka. Je neuróny majú inhibičný vplyv na cieľové štruktúry. Funkčne patrí do bazálnych ganglií, ale morfologicky je súčasťou medzimozgu. Na horizontálnom reze cez bazálne gangliá je možné popísať nasledovné štruktúry (z laterálnej strany mediálnym smerom).
- insula
- capsula extrema
- claustrum
- capsula externa
- putamen
- lamina medullaris lateralis
- globus pallidus
- crus posterius capsulae internae – thalamus
- crus anterius capsulae internae – ncl. caudatus
Čierna hmota – substantia nigra
Substantia nigra je umiestnená v celom rozsahu stredného mozgu ako jeho najväčšie neurotransmiterov v nigro-striatových prepojeniach. Funkčne je substantia nigra zapojená do bazálnych ganglií, ale morfologicky sa radí k strednému mozgu. Anatomicky sa k bazálnym gangliám zaraďujú aj mandľovité teleso, corpus amygdaloideum (amygdala) a závora, claustrum.
Spojenia bazálnych ganglií
Spojenia bazálnych ganglií sú nesmierne zložité, zjednodušené, na opisné účely, sa delia na tri základné typy:
- Aferentné – prívodné vlákna prichádzajú predovšetkým z kôry mozgu. Menšia časť informácií prichádza aj z interlaminárnych jadier lôžka.
- Vzájomné prepojenia medzi jednotlivými jadrami bazálnych ganglií:
- 1) dôležité striatopallidové spojenia
- 2) recipročné spojenia medzi striatum a substantia nigra
- 3) recipročné spojenia medzi pallidum a ncl. subthalamicus
- Eferentné – výstupné vlákna vychádzajú hlavne z pallida a zo substantia nigra a smerujú do lôžka, do ncl. ventralis anterior. Z lôžka vystupujú lôžkovokôrové vlákna do kôry mozgu.
Funkcia bazálnych ganglií
- Bazálne gangliá sú zapojené do riadenia motoriky ako koordinačný činiteľ medzi neúmyselnou reflexnou činnosťou a úmyselnými pohybmi. Vývinovo staršie pallidum sa zúčastňuje na riadení svalového napätia, kým mladšie striatum súvisí s riadením celkových, rozsiahlých pohybov. Základnou funkciou bazálnych ganglií je predovšetkým riadenie zložitých vzťahov medzi podráždením a útlmom pri úmyselných pohyboch. Všeobecným rysom ich činnosti je tlmivý vplyv na kôrové a podkôrové motorické funkcie. Aktivitu primárnych motorických oblastí upravujú buď priamym pôsobením na neuróny mozgovej kôry alebo tlmia kôrovú výstupnú informáciu na úrovni nižších oblastí CNS , predovšetkým retikulárnej formácie a miechových reflexov. V oboch prípadoch teda korigujú informáciu z primárnej motorickej oblasti skôr, než dôjde k motoneurónom predných rohov miechy, t.j. ku každej spoločnej dráhe. Výskumy ukázali, že neuróny striata zvyšujú svoju vzruchovú aktivitu už pred začiatkom pohybu, z čoho vyplýva, že bazálne gangliá sa zúčastňujú na programovaní pomalých a ustálených pohybov. Tlmivú činnosť bazálnych ganglií predstavujú okrem spojení s motorickými oblasťami mozgu aj spojenia zo substantia nigra a s retikulárnou formáciou (s inhibičnou oblasťou).
Stručne možno charakterizovať funkciu bazálnych ganglií okruhom: kôra mozgu – striatum – palllidum – thalamus – kôra mozgu. V súčasnosti je tento okruh niekoľkých okruhov, v ktorých sú zapojené rozdielne časti bazálnych ganglií, rôzne jadrá lôžka a rozdielne oblasti mozgovej kôry.
Podľa zapojených štruktúr sa rozoznávajú:
- motorický okruh –začína v senzitívnomotorickej kôrovej oblasti,
- limbický okruh – vystupuje z gyrus cinguli, amygdaly a hipokampu
- kognitívny okruh – vychádza z prefrontálnej kôry
- osobnostný okruh – vystupuje z orbitofrontálnej kôry (kôra čelového laloka v oblasti gyri orbitales).
Čuchový mozog – rhinencephalon
Ak uvádza Kluchová (2015) najvýznamnejšie štruktúry čuchového mozgu sú čuchová guľa, bulbus olfactorius, prístredný čuchový prúžok, stria olfactoria medialis, bočný čuchový prúžok, stria olfactoria lateralis a primárna čuchová kôra, area piriformis. Čuchové štruktúry sú dobre vyvinuté u nižších stavovcov, u ľudí tvoria len malú časť mozgu. Termín rhinencephalon (čuchový mozog) sa ôvodne používal pre mnoho štruktúr limbického systému, v súčasnosti sa považuje za vhodné označovať takto len štruktúry priamo spojené s čuchovou funkciou.
Čuchové štruktúry
Po podráždení čuchových receptorových neurónov v čuchovej sliznici (1.neurón) prechádza nervový vzruch axónom cez riečicovitú (dierkovanú) platňu , lamina cribrosa, do čuchovej gule, bulbus olfactorius, v ktorej sa pripojí na mitrálne bunky (2. neurón). Ich axóny sú dlhé, vstupujú do čuchovej dráhy, tractus olfactiorius, ktorá sa smerom dozadu delí na prístredný čuchový prúžok, stria olfactoria medialis a bočný čuchový prúžok, stria olfactoria lateralis. Oba prúžky tvoria prednú hranicu prednej dierkovanej hmoty, substantia perforata anterior. Cez stria olfactoria medialis prechádzajú čuchové vlákna do druhostranného tractus olfactorius a do oblastí priehradky, septum. Väčšia časť čuchových vlákien však prechádza cez stria olfactoria lateralis a smeruje do oblasti primárnej čuchovej kôry, area piriformis. Je to ontogeneticky stará oblasť. Za sekundárnu čuchovú kôru sa považuje entorinálna oblasť, area entorhinalis, ktorá leží rostrálnej a laterálnej časti uncus gyri parahippocampalis. Primárna a sekundárna čuchová kôra tvoria spolu čuchový lalok, tzv. hruškovitý lalok, lobus piriformis.
Čuchové dráhy
Čuchový systém je výnimočný v tom, že ako jediný vysiela vlákna neurónov priamo do kôry mozgu, bez prepojenia v lôžku. Ďalšie spojenia vedú do orbitofrontálnej kôry, kde dochádza k analýze čuchových vnemov. Významne spojenia primárnej čuchovej kôry smerujú do:
- lôžka – do ncl. dorsomedialis a odtiaľ do orbitofrontálnej kôry, čo zaisťuje vedomé vnímanie čuchovej informácie.
- entorinálnej oblasti (laterálna časť uncus gyri parahippocampalis)– kde sa spracuvávajú pamäťové procesy spojené s čuchom
- amygdaly (v hĺbke uncus gyri parahippocampalis) – táto štruktúra je významne zapojená do limbického systému, realizuje spojenie čuchových vnemov s emočnými reakciami. Amygdala teda sprostredkuváva emocionálne podfarbenie čuchových vnemov. Ďalšie spojenia vedú do orbitofrontálnej kôry, kde dochádza k analýze čuchových vnemov. Čuchový systém má veľký význam v spojení s limbických systémom, považuje sa za vysunuté tykadlo limbického systému.
Limbický systém – systema limbicum
Kľúčovú úlohu na fungovaní limbického systému má hipokampus, amygdala a podlôžko. Termínom limbický systém sa označuje funkčný systém kôrových a podkôrových neurónov. Spojenia medzi nimi tvoria komplexné okruhy, ktoré majú dôležitú úlohu pri získavaní a udržiavaní pamäti, pri emóciách a správaní.
Všeobecná charakteristika
Opísať neurofyziologický základ dejov v limbickom systéme je náročné, pretože doterajšie znalosti o neurónových mechanizmoch emočných reakcií stále nie sú úplné. Experimentálne štúdie a klinické pozorovania ukazujú, že hlavnou riadiacou oblasťou týchto dejov je jeden z najzložitejších integračných systémov mozgu – limbický systém. Zložitosť limbického systému vyplýva z jeho vývinu. V raných fázach embrynálneho vývinu, sú hipokampus, septum, amygdala aj podlôžko uložené tesne vedľa seba. Dráhy, ktorými sa navzájom prepájajú, sú preto priame a krátke. S vývinom predného mozgu sa však časti limbického systému od seba vzdialili a vznikla zložitá organizácia často veľmi dlhých vnútorných spojení. Názov limbický súvisí s morfologickým usporiadaním jeho kôrovej časti ktorá počas vývinu vytvorila akýsi golier, hranicu (limbus) okolo mozgového kmeňa. K limbickému systému patria niektoré štruktúry mozgovej kôry (prefrontálna kôra, septum, gyrus cinguli, gyrus parahippocampalis, hipokampová formácia) a niektoré podkôrové oblasti (amygdala, lôžko, podlôžko, stredný mozog). Kôra mozgu, cortex cerebri, sa z vývinove hľadiska delí na tri rôzne oblasti, v každej z nich sa nachádzajú štruktúry, patriace k limbickému systému. K podkôrovým oblastiam limbického systému patrí hlavne amygdala. V širšom význame sem patria aj niektoré jadrá lôžka, podlôžka, nadlôžka a retikulárnej formácie.
Najvýznamnejšie štruktúry limbického systému
K štruktúram limbického systému patria lobus limbicus, amygdala, hipokampová formácia a hypothalamus. Medzi najvýznamnejšie z nich (limbický systém v užšom zmysle) patria hipokampová formácia, corpus amygdaloideum a septum verum. Lobus limbicus však neznamená to isté čo limbický systém, pretože plní aj iné ako nečuchové funkcie.
Hipokampová formácia
Zo všetkých zložiek limbického systému je hipokampová formácia jediná, ktorá je výlučne jeho súčasťou. Ostatné zložky limbického systému sú zároveň súčasťou iných neuroanatomických štruktúr a systémov (napr. corpus amygdaloideum je zapojené j do čuchového systému, septum verum súvisí aj s autonómnymi funkciami podlôžka).
– pozostáva z troch štruktúr: vlastný hippocampus, gyrus dentatus a subiculum.
- Hippocampus– bol počas vývinu vytvorený vtlačením a zatočením spodnej časti spánkového laloka do bočnej mozgovej komory, Nachádza sa na spodine dolného rohu bočnej mozgovej komory. Na priečnom reze má tvar morského koníka.
- Gyrus dentatus– je zúbkovitá struktúra, uložená pozdĺž mediálneho okraja hipokamu.
- Subiculum (podlôžka)– je úzky pruh kôry uložený v sulcus hippocampi.
- Funkciia hipokampovej formácie vyplýva z jej účasti pri vytváraní pamäťových stôp, pri učení (bilaterálnej) temporálnej lobektómii pri epilepsi) dochádza k amnézii (strate pamäti) pacienta. Základné charakteristiky takejto amnézie sú:
- udalosti pred poškodením hipokampu sú pamäťovo uchované,
- dlhodobá pamäť sa uchováva
- kratkodobá pamäť je uchovaná len na niekoľko minút
- Aferentné vlákna k hipokampu prichádzajú z: gyrus cinguli, septálne oblasti (cez fornix), entorinálnej oblasti.
- Eferentné vlákna z hipokampu vedú do: septa (cez fornix), podlôžka a corpus mammillare (cez fornix), ncl. anterior thalami (cez fornix), mozgového kmeňa (tegmentum stredného mozgu, cez FTM),habenulárnych jadier (nepriamo cez stria medullaris thalami).
- Hlavnou eferentnou dráhou hipokampu je fornix.
Mandľovité teleso – corpus amygdaloideum (amygdala)
– má mandľovitý tvar a je uložená v rostrálnej časti spánkového laloka tesne pred hipokampom. Amygdala je významná pri tvorbe negatívnych emócií a s tým spojených somatických motorických reakcií, vrátane reakcií orgánov. Po jej podráždení sa u ľudí dostavia zúrivé a agresívne reakcie, nazýva sa aj centrum agresivity. Odstránenie amygdaly má za následok poslušné a flegmatické správanie, stráca sa schopnosť emočného sprievodu správania a emočnej formy komunikácie. Amygdala funguje aj ak emočná zložka pri bolesti. Z pohľadu tvorby emócií je významné aj spojenie amygdaly so septom. Vďaka svojmu zapojeniu medzi mozgovou kôrou a ostatnými časťami limbického systému, predstavuje amygdala okno, ktorým limbický systém pozoruje človeka v okolitom svete.
Aferentné dráhy k amygdale prichádza z: čuchovej, limbickej a asociačnej kôry, podlôžka (hypothalamus).
Eferentné dráhy z amygdaly smerujú do: septa. cez stria terminalis a stria diagonalis), podlôžka ( cez ventrálnu amygdalofugálnu dráhu – VAD a stria terminalis), ncl. mediodorsalis lôžka, mozgovej kôry (limbická a asociačná).
Hlavné eferentné dráhy amygdaly sú: stria terminalis (spája amygdalu s podlôžkom s septom), ventrálna amygdalofugálna dráha (spája amygdalu s podlôžkom, lôžkom – ncl. dorsomedialis a mozgovým kmeňom).
Septum – septum verum
– je lokalizované pod rostrálnou časťou corpus callosum, je to kôrová oblasť uložená pred lamina terminalis a oblasť v okolí commissura anterior. Najdôležitejšie štruktúry septum verum sú gyrus paraterminalis a area subcallosa. Nad septum verum sa nachádza septum pellucidum (medzi fornix a corpus callosum). Funkciou septa je jeho naviazanie na tvorbu pocitov radosti a odmeny a emočných stavov vyvoláných po požití drog, ako sú nikotín, marihuana, kokaín a heroín, ale aj postkoitálne pocity. Oblasť septa sa nazýva aj ako centrum príjemností. Septum je prepojené aj s amygdalou, z čoho vyplýva ich úzka súčinnosť pri tvorbe emócií. Štruktúry septa produkujú 5-hydroxytryptamín (serotonín), jeho nedostatok sa u pacienta prejaví psychickými zmenami (depresie).
Aferentné dráhy k septu prichádzajú z: amygdaly (cez stria diagonalis a stria terminalis), hipokampovej formácie (cez fornix)
Eferentné dráhy septa: septum je súčasťou multisynaptickej dráhy fasciculus telencephalicus medialis ( FTM), ktorá spája kôru mozgu s mozgovým kmeňom: prostredníctvom tejto dráhy je septum priamo spojené s podlôžkom, významnou dráhou zo septa je aj stria medullaris thalami, ktorá predstavuje spojenie s habenulárnymi jadrami a pokračuje ďalej až do mozgového kmeňa (spojenie s noradrenergickými jadrami).
Hlavné eferentné dráhy septa sú: FTM septum – hypothalamus (ďalej pokračovanie do stredného mozgu), stria medullaris thalami: septum – ncll. habenulares (a ďalej do stredného mozgu).
Hipokampus, amygdala aj septum sú priamo spojené s podlôžkom (hypothalamus). Odtiaľ je realizované spojenie s autonómnymi jadrami mozgového kmeňa a miechy (cez fasciculus longitudinalis posterior).
Spojenia limbického systému
Vstup informácií (inštinktívnych vnemov) do limbického systému môže prebehnúť dvojakou cestou: priamo, cez amygdalu, ktorá poskytuje emocionálne vnímanie daného impulzu, obzvlášť v spojení so sociálnym významom. V evolučnom vývoji zohráva amygdala dôležitú úlohu pri vyvažovaní primitívneho a racionálneho správania. Reakcie na primitívne vnemy sprostredkúva autonómny nervový systém, vychádzajúci z podlôžka.
Nepriamo cez hipokampovú formáciu, prostredníctvom area entorhinalis, ktorá zabezpečuje spojenie daného impulzu s predošlou skúsenosťou, pretože hipokampus zohráva úlohu v procese učenia a pamäti.
Nervové dráhy zapojené do limbického systému
Fornix tvoria vlákna, ktoré smerujú z hipokampu do podlôžka (corpus mammillare), lôžka a septálnej oblasti, ako aj zo septa späť do hipokampu. Fornix tvoria dva útvary. Hipokampus pokrýva jemný poprašok, alveus, ktorý je vlastne tenkou vrstvou bielej hmoty. Sú to nervové vlákna zbiehajúce sa na mediálnom okraji hipokampu, kde vytvárajú strapec, fimbria hippocampi. Fimbria smeruje dozadu a pod splenium corporis callosi opúšťa hipokampus ako rameno oblúka, crus fornicis, ktoré sa stáča smerom nahor a prepája sa s druhostraným crus fornicis. Obe časti crus fornicis sú prepojené priečnymi vláknami – spojkou oblúka, commissura fornicis. Tieto vlákna sa krížia a zabezpečujú tým vzájomné spojenie oboch hipokampov (z obidvoch spánkových lalokov). V ďalšom priebehu crus fornicis zhrubne, nazývame ho telo oblúka, corpus fornicis. Telá oboch oblúkov, približené tesne k sebe, prechádzajú nad lôžkom a tesne pod corpus callosum. Corpus fornicis smeruje dopredu a nad predným okrajom lôžka sa rozdeľuje do dvoch stĺpcov, columnae fornicis, ktoré sa stáčajú smerom nadol a laterálne, zanárajú sa do spodiny III. mozgovej komory (do podlôžka) a končia v bradavkovitom telese, corpus mammillare. Cingulum slúži ako asociačný zväzok pre priľahlé oblasti mozgovej kôry v čelových, temenných, záhlavných lalokoch a v lobus limbicus. Spája tiež oblasť septa s gyrus parahippocampalis. Ventrálna amygdalofugálna dráha (VAD), tractus amygdalohypothalamicus, vedie z amygdaly do podlôžka, do lôžka, mozgového kmeňa a miechy. Stria diagonalis – Broca sa nachádza tesne za tractus opticus a tvorí zadný okraj substantia perforata anterior. Funkčne ide o spojenie amygdaly so septálnou oblasťou. Commissura anterior je zväzok vlákien idúci priečne za lamina terminalis. Vzájomne prepája bulbi olfactorii, predné časti spánkových lalokov a obe amygdaly. Tractus mammillothalamicus poskytuje dôležité spojenie medzi corpus mammillare a ncl. anterior thalami. Tractus mammillotegmentalis spája corpus mammillare a stredný mozog. Stria medullaris thalami tvorí biely pruh na rozhraní medzi mediálnou a dorzálnou plochou lôžka a vedie vlákna zo septa do oblasti ncll. habenulares. Stria terminalis je uložená medzi thalamus a ncl. caudautus, pričom sleduje zakrivenie ncl. caudatus po jeho mediálnej strane. Vlákna stria terminalis vychádzajú z amygdaly a smerujú do podlôžka a septa. Fasciculus telencephalicus medialis (FTM) – je významným spojením oblasti septa s podlôžkom a ďalej až s mozgovým kmeňom. Facsiculus longitudinalis posterior (FLP) – spája podlôžko s autonómnymi centrami mozgového kmeňa. Je zodpovedný za autonómne reakcie pri emočných stavoch, a to tak, že prvotná je informácia z amygdaly do podlôžka a ž potom sa spomenutým FLP dostaví autonómna odpoveď. Stria olfactoria medialis et lateralis čuchového mozgu.
Funkcia limbického systému
Impulz z mozgovej kôry (neocortex) prichádza do limbického systému, ktorý spája uvedomelé myslenie s primitívnym inštiktívnym cítením a s vnútornou homeostázou prostredníctvom mnohých nervových spojení. Zjednodušene je to možné vyjadriť tak, že informácie z vonkajšieho sveta, prichádzajú špecifickými dráhami (napr. zrak, sluch, dotyk), sú filtrované v temennozáhlavovej kôre mozgu. Odtiaľ sú tieto informácie prenášané do čelovej kôry, v ktorej dochádza k regulácií správania a tiež do spánkovej kôry, kde sa uvedené informácie významovo spracúvajú. Až potom sa kôrové impulzy dostávajú do limbického systému, kde sa spájajú s inštinktívnymi vnemami.
Všeobecne možno zhrnúť funkciu limbického systému do troch kategórií:
- somatické motorické odpovede: patria sem reakcie týkajúce sa jedenia, pitia, sexuálnych funkcií
- autonómne odpovede: dráhy idúce z limbického systému sú vo veľkej miere sprostredkované spojeniami s podlôžkom. S tým súvisia aj zmenené funkcie orgánov (kardiovaskulárne, gastrointenstinálne a pod. )
- senzorické odpovede: limbický systém sprostredkúva mnoho pocitov súvisiacich s emóciami, či už pozitívnymi alebo negatívnymi. Sem patrí aj vnímanie čuchových stimulov a viscerálnych (orgánových) podnetov.
Pre rámcové funkcie limbického systému je charakterizovaný tzv. Papezov okruh, ktorý poskytuje štrukturálny rámec pre vyjadrenie emócií. Papezov okruh tvorí jednoduché spojenia, ktoré boli automaticky potvrdené. Uvedený okruh je charakterizovaný takto: hippocampus – fornix – corpus mammillare – ncl. anterior thalami – gyrus cinguli – hippocampus.
Emócie
Ako uvádza Kluchová (2015) emócie sú spojené s aktiváciou štruktúr retikulárnej formácie, limbického systému a medzimozgu. Hlavným centrom emócií je limbický systém. Emóciou sa najčastejšie označuje citové zafarbenie činnosti alebo psychický stav, ktoré majú vplyv na správanie jedinca. Emócie sú vyjadrované výrazom tváre alebo aj charakterickým postojom tela a polohou jednotlivých jeho častí (tzv. neverbálna komunikácia). Výraz tváre je tvorený súhrou mimických svalov, ktoré sú ovládané prostredníctvom tvárového nervu.
Čo sú to emócie
Pojem emócie doslovne znamená citové rozpoloženie. Emócie zohrávajú dôležitú úlohu v reakciách a rozhodnutiach jednotlivca. I keď sa emócie vyskytujú aj u iných druhov, existuje niečo celkom zvláštne v spôsobe, akým sa emócie stávajú komplexom myšlienok, princípov a názorov, ktoré sa vyskytujú len u nás, ľudí. Emócie je možné posudzovať ako: subjektívny fenomén – emocionálne prežívanie, ktoré patrí do psychologickej sféry, objektívny fenomén – emočné správanie, prístupné aj pozorovaniu. Emočné správanie je sprevádzané aj zmenami v somatickej a viscerálnej oblasti (napr. smiech, plač, grimasy, zmeny krvného tlaku, frekvencie srdca, peristaltiky čriev a pod.)
Emócie predstavujú sériu mechanizmov, ktoré sa rozvinú za účelom: vyrovnania sa s ohrozením alebo nebezpečím – odčerpávací systém, napr. môžeme rozvinúť bojovú odozvu voči ohrozeniu, keď sa krv nahrnie k hornej časti tela, k ramenám a rukám, alebo únikovú odozvu, keď sa prekrví mohutné svalstvo nôh – v oboch prípadoch ide o reakcie spojené s autonómnym nervovým systémom/prípravy k dejom, ktoré môžu byť užitočné nám alebo našim génom – prístupový systém, napr. môžeme byť motivovaní zabezpečiť si životné potreby alebo partnera a uspokojenie týchto potrieb sprevádzajú pocity radosti a odmeny. Pri emóciách nepracuje mozog nezávisle od tela, ale v súčinnosti s telom. Všetky emócie sú sprevádzané telesnými pocitmi, vyjadreniami alebo odozvami.
Čo emócie spôsobujú
– poskytujú motiváciu pre kritické konanie a prejavy a to: v podnetnom prostredí (súťaživosť – prístupový systém), pri averzívnych stimuloch(bojovo–únikové konanie – odčerpávací systém), prispôsobujú pamäť, ovplyvňujú schopnosť učenia a urýchlujú proces rozhodovania, zohrávajú kľučovú úlohu pri rozdielnostiach medzi osobnosťami a pri stupni zraniteľnosti, ktorý ovplyvňuje vznik psychických porúch.
Klasifikácia ľudských emócií
Emócie sú mimovoľné, neúmyselné, na rozdiel od myšlienok, ktoré sú väčšinou dobrovoľné a súkromné. Môžeme sa uvážene rozhodnúť, o čom chceme rozmýšľať, ale nemôžeme sa rozhodnúť, či budeme mať emóciu, či nie.
Existuje sedem základných ľudských emócií, ktoré boli zistené vo všetkých študovaných kultúrach ľudí: strach, hnev, odpor, smútok, šťastie, prekvapenie a opovrhovanie.
Neurálny podklad emócií
V posledných rokoch sa objavili pokusy objasniť úlohu nervového systému vo vzťahu k emóciám. Vo všeobecnosti je možné konštatovať, že najvýznamnejšie miesta, z ktorých sa vyvolávajú pozitívne emócie, sú v špecifických častiach limbického systému a vo ventromediálnej časti podlôžka. Naopak, negatívne emócie možno vyvolať dráždením jeho dorzolaterálnej časti a časti stredného mozgu. Je zrejmé, že u ľudí existuje značná individuálna variabilita v časovom priebehu reakcie na emócie, v intenzite tejto odozvy a tiež v dobe návratu do pôvodneho stavu. Spracovávanie podnetov pomocou mozgových štruktúr na odlišných úrovniach prispieva k emocionálnemu prežívaniu rozličným spôsobom. Bazálne, pudové reakcie sú riadené podkôrovými oblasťami. Na úrovni limbického systému sú emocionálne reakcie tohto pudového systému modulované a vznikajú tu primárne emócie s ich mimickými prejavmi. Kôrové oblasti mozgu umožňujú uvedomiť si tieto emócie a vo väčšine prípadov ich určitým spôsobom modelovať a kontrolovať.
Úloha amygdaly
Pri študovaní podkôrových štruktúr, ako reprezentantov emócií, sa ukázala byť významnou amygdala, ktorá je možné charakterizovať ako centrum nepríjemných pocitov.
- pokusy na zvieratách potvrdili účasť amygdaly pri strachu a pri reakcii na strach,
- výskumy u ľudí, ukázali, že amygdala zohráva rozhodujúcu úlohu pri spracovaní negatívnych emócií (porušená schopnosť rozoznať výraz strachu v tvári, posudzovanie neznámych ako nedôveryhodných
- zobrazovacie techniky potvrdili aktiváciu amygdaly pri odpovedi na strach, v spoločnosti nepríjemných osôb, pri určení nepríjemných úloh a pri odporných chuťových a čuchových podnetov
Podľa výsledkov výskumov sa možno domnievať, že amygdala zohráva kľučovú úlohu pri štúdiu nových, nepredvídateľných podnetov, hrozieb. U ľudí sa pozorovali individuálne rozdiely v metabolickej aktivite amygdaly. Potvrdili sa rozdiely u jednotlivcov v glukózovom metabolizme amygdaly pri nepríjemných podnetoch.
Správanie dieťaťa v prenatálnom období
Ako uvádzajú Porter et al. (1997) in Langmeier et. Krejčířová (2006) plod je veľmi skoro pripravovaný pre činnosti, ktoré budú potrebné pre jeho prežitie a pre interakciu s vonkajším svetom po narodení. Už na konci druhého mesiaca, keď embryo dosiahlo dĺžku 25 mm a kde sa vytvoril základ všetkých svalových skupín, je možné pozorovať reakcie vo forme jemných kontrakcií, aj keď svalová sústava ešte nie je napojená na nervový systém. Na začiatku fetálneho vývinu, začiatkom tretieho mesiaca, plod už spontánne pohybuje hornými a dolnými končatinami, otáča hlavičku, zvrášťuje čelo, otvára a zatvára ústa a čoskoro sa ukazujú náznaky úchopových pohybov (aj keď matka pohyby ešte dlho nevníma). V deviatich týždňoch je možné už možné registrovať bioelektrickú aktivitu mozgu na EEG, najprv nerozlíšenú, od piateho mesiaca sa na zázname začína líšiť postupne spánkový a bdelý stav. Plod reaguje aj na podnety vizuálne (svetelné lúče dopadajúce na brušnú stenu matky), zvukové, na tlak i na bolesť. Senzorické dráhy bolesti fungujú od 20. týždňa, aj keď nie je doposiaľ známe, či veľmi malé deti vnímajú bolesť rovnako intenzívne ako dospelí jedinci. Je však známe, že nedonesené deti sú na bolesť veľmi citlivé. Schopnosť habituácie na zvuk a na vibrácie je preukázaná od 22. týždňa tehotenstva a s vekom narastá. Túto schopnosť ide teda dobre použiť ku zisťovaniu schopnosti vnímať a rozlišovať rôzne podnety plodom. Plod je napríklad schopný rozlíšiť slabiky ľudskej reči (bibi od baba), v 35. týždni tehotenstva. Schopnosť habituácie pritom závisí na aktuálnom stave CNS a zhoršuje sa napríklad pri nedostatku kyslíka u matky, pri podávaní liekov matke (napr. Diazepam), ale i do 1,5 hodiny od vyfajčenia jednej cigarety, či dokonca pri emočnom strese matky. Abnormálnu habituáciu nachádzame samozrejme tiež u detí s vrodenými vývinovým vadami CNS alebo u detí s genetickým postihnutím. Ako už preukázali staršie štúdie, že plod je v posledných dvoch mesiacoch vnútromaternicového života už schopné sa učiť, zbierať skúsenosti. Okrem uvedených habituačných reakcií je preukázaná schopnosť dieťaťa vytvárať podmienené spojenie medzi zvukom a podnetom. Plod sa tak zdokonaľuje pre adaptáciu na svet, ktorý naňho po narodení čaká, a s ktorými sa musí vyrovnávať , rovnako ako s pocitmi, vyvolanými biologickými podnetmi z vlastného tela. Plod je aktívny, tzn. ovláda, kontroluje prostredie. Už v prenatálnom období získava schopnosť jedinec aktívne sa zúčastňovať interakcie s fyzikálnym prostredím. Čoskoro sa jedince dostáva do určitého aktívneho kontaktu s matkou. Na jeho spontánny pohyb matka emocionálne reaguje a jej emócie opäť ovplyvňujú dieťa vytváraním vzorcov zvláštnych podnetových situácií. Rhigetti (2000) opisuje v empirickej štúdií, že emočné zmeny matky (keď matka sleduje relaxačné alebo naopak aktivačné podnety) vedú v 30. týždni tehotenstva u dieťaťa ku zmenám tepu a spontánnym pohybom. Dieťa je vtedy s matkou už emočne vyladené. Takto sa ešte pred narodením vytvára s matkou určitý dialóg, ktorý sa po narodení rozvíja v zmysluplnú neverbálnu komunikáciu. Ako autori Matějček et. Langmeirer (1986) in Langmeier et. Krajčířová (2006) uvádzajú, dobrý psychický stav matky je pre základy psychiky dieťaťa veľmi dôležitý.
Bdelý stav a spánok
Ako uvádza Králíček (2023) zvláštnou zložkou nervovej činnosti je stav, ktorý označujeme ako vedomie. Vedomia má dvojitý význam, opisuje vedomie seba samého a schopnosť vnímať a prežívať realitu okolitého sveta. Takto porozumené vedomie sa označuje ako vigilita. V polovici 30. rokov 20. storočia belgický neurofyziológ Frederic Bremer ukázal, že preťatie mozgového kmeňa medzi colliculi superiores a inferiores – vyvolá, že zviera upadá do bezvedomia a elektrická aktivita mozgu je charakterizovaná spánkovým typom elektroencefalogramu. Oproti tomu preťatie miechy nad prvým miechových segmentom a zaistenie dýchania riadenou ventiláciou – spôsobí, že zviera strieda normálny spánok a bdenie a keď je v bdelom stave, elektrická aktivita mozgu charakterizuje normálny bdelý stav. Uvedené pokusy ukázali, že v mozgovom kmeni existujú určité nervové štruktúry dôležité pre udržanie bdelého stavu.
Spánok
Spánok môžeme definovať ako stav bezvedomia, z ktorého človek môže byť prebudení primeraným senzorickým podnetom. Spánok ide pomerne dobre charakterizovať elektroencefalograficky. Pri EEG meraniach sa zistilo, že spánok nie je jednotný, ale že existujú dva spánkové stavy:
- non– REM (NREM) spánok alebo tiež označovaný ako spánok s pomalými vlnami
- REM spánok alebo tiež spánok paradoxný
Následky deprivácie REM spánku – pri monitorovaní EEG, elektrookulogramu a elektromyografickej aktivity v priebehu noci ide identifikovať periódy REM spánku a jej ďalší priebeh zrušiť prebudením pokusnej osoby. Pôvodne sa uvažovalo, že vylúčenie typu REM spánku môže viesť k vážnym psychickým poruchám. Táto hypotéza však nebola potvrdená. Na druhej strane sa však objavil zaujímavý fakt, že odstránením REM spánku po obdobie niekoľko nocí spôsobuje, že pri nasledujúcom neprerušovanom spánkusa celkový počet a trvanie jednotlivých periód REM spánku výrazne zvýši. Ako by chcel organizmus vykompenzovať predošlú stratu tohto spánkového stavu.
Neurofyziologické mechanizmy vzniku spánku
V 50. rokoch 20. storočia prevládala predstava, že spánok je pasívny stav, navodený poklesom bdelosti pri obmedzenej senzorickej stimulácií mozgu. V dôsledku toho sa výskum nervových mechanizmom zameral na tie nervové systémy, ktoré udržujú stav bdelosti. V 60. rokoch sa však táto koncepcia radikálne zmenila. Spánok začal byť za aktívny neurofyziologický proces, navodený aktivitou iných ústredí centrálneho nervstva. Je známe, že mechanizmy generujúce non – REM a REM typu spánku sa nachádzajú v anatomicky odlišných častiach CNS. non– REM spánok je kontrolovaný komplexným systémom, ktorého kľúčové neuronálne zoskupenie sa nachádzajú v nasledujúcich oblastiach mozgu:
najkaudálnejšia oblasť, ktorá má vzťah k non– REM spánku, je nucleus tractus solitari. Ide o jadro predĺženej miechy , nachádzajúci sa pozdĺž tractus solitarius, bolo preukázané, že jeho nízkofrekvenčná elektrická stimulácia navodzuje non– REM spánok
nuclei raphe – ide o jadrové zoskupenie serotonergných neurónov retikulárnej formácie predĺženej miechy, jeho rozrušenie alebo farmakologická blogáda syntézy serotonínu pomocou p– chlorfenylalanínu spôsobí nespavosť
cholinergné neuróny retikulárnej formácie pontu a mesencephala
nucleus reticularis thalami – ide o nešpecifické jadro thalamu, predpokladá sa, že generuje spánkové vretená v EEG zázname
štruktúry pars basalis telencephali– patria sem jadrá diagonálneho zväzku – ncl. basalis (Meynerti), substantia innominata a laterálna preoptická oblasť hypothalamu. Elektrická stimulácia týchto štruktúr môže navodiť non– REM spánok. Zaťial čo non– REM spánok je kontrolovaný rozsiahlou neuronálnou sieťou, zdá sa, že REM spánok je generovaný diskrétnou oblasťou v nucleus reticularis pontis oralis. Ide o jadrové zoskupenie v rostrálnej časti ponton retikulárnej formácie. Jeho rozrušenie permanentne eliminuje REM spánok.
Cyklické striedanie spánku a bdenia
Takmer u všetkých rastlín a živočíchov sa stretávame s cyklickými zmenami mnoho funkcií. Tieto cykly sú rôzne dlhé. Najvýraznejší je denný (diurnálny, cirkadiánni) rytmus, ktorý trvá 24 hodín. Dennému rytmu podlieha aj cyklické striedanie spánku a bdenia. V súčasnej dobe sa zisťuje, že biologické hodiny – určujúce striedanie spánku a bdenia ležia v hypothalame, a to v nucleus suprachiasmaticus. Rozrušenie tohto jadra celko rozvráti cyklus bdenia a spánku. Bolo zistené, že vnútorný pacemaker je teda nastavený na 25 hodín. Ďalej bolo zistené, že charakteristické denné striedanie svetla a tmy pôsobí na vnútorný cirkadiánny pacemaker prostredníctvom retinohypothalamických spojov, ktoré prinášajú informácie o osvetlení sietnice priamo do ncl. suprachiasmaticus.
Psychické poruchy, osobnosť jedinca a emócie
Pacienti s poruchami psychiky predstavujú problém pre svoje okolie aj pre seba. Takíto ľudia môžu byť agresívni a škriepni alebo zákerní a môžu sa správať manipulatívne a nepredvídateľne. Psychické poruchy vznikajú vzájomnou súčinnosťou povahových faktorov a raných, skorých skúseností z prostredia (nature and nurture). Je dôležité rozoznať, kedy správanie jedinca už nie je fyziologické, ale poukazuje na psychickú poruchu. Psychickú poruchu je možné rozoznať od: normálneho štýlu, etapy návratu reakcií do normálu, anxióznych ochorení.
O PSYCHICKEJ PORUCHE JE POTREBNÉ UVAŽOVAŤ , KEĎ PACIENTOVA CHARAKTERISTICKÁ KONŠTELÁCIA EMÓCIÍ, MYŠLIENOK, MEDZIĽUDSKÝCH VZŤAHOV NARÚŠA JEHO SCHOPNOSŤ MILOVAŤ, PRACOVAŤ A ZABÁVAŤ SA.
Osobnosť a mozog
Medzi odborníkmi sa často diskutuje o otázke medzi rozlišovaním medzi osobnosťou a povahou.
POVAHA – je biologická danosť, je to vnútorná predispozícia, ktorá určuje jedincovi, ako sa zachová, keď má čeliť rôznym sociálnym, emocionálnym a poznávacím podnetom.
OSOBNOSŤ – je jedinečný súbor vlastností a skúseností – súhra povahy a výchovy, ktorá podfarbuje spôsob, akým jedinec reaguje na okolitý svet. Pri stanovovaní charakteristiky osobnosti sa 50– 70% negatívnych prejavov pripisuje dedičným faktorom.
Zoznam použitej literatúry
KRÁLÍČEK, P. (2023). Úvod do speciální neurofyziologie. Praha: Galén, 235 s. ISBN 978-80-7492-641-9.
LANGMEIER, J. – KREJČÍŘOVÁ,D. (2006). Vývojová psychologie. Praha: Grada, 368 s. ISBN 978-80-247-1284-0.
LOVÁSOVÁ, K. – KLUCHOVÁ, D. – BOLEKOVÁ, A. (2015). Neuroanatómia pre psychológov, štruktúry a funkcie nervového systému. Univerzita Pavla Jozefa Šafárika v Košiciach, Filozofická fakulta, 219 s. ISBN 978-80-8152-231-4.
PROCHÁZKA, R. – OREL, M. (2021). Vývojová neuropsychologie. Praha: Grada, 223 s. ISBN 978-80-271-3080-1.